Revista Biomédica Revisada Por Pares

 Para Descargar PDF debe Abrir sesión.
Para Descargar PDF debe Abrir sesión.
Palabras clave: fatty liver, liver regeneration
La esteatosis hepática es el acúmulo de grasa en los hepatocitos, que puede ser la consecuencia del proceso de regeneración hepática o de procesos patológicos como la enfermedad del hígado graso alcohólica y no-alcohólica. A pesar de su importancia, en ambos casos el mecanismo exacto en que la esteatosis influye en la regeneración hepática es escasamente comprendido. Estudios previos han demostrado que los pacientes que presentan hígado graso expresan una regeneración hepática dispar, posiblemente debido a la acumulación de especies reactivas de oxígeno generada por procesos inflamatorios ocasionados por activación de las células de Kupffer. En este artículo se revisan distintos factores que afectan la regeneración hepática, tratando de comprender el mecanismo involucrado en la regeneración dispar en hígados con esteatosis ocasionada por una enfermedad de hígado graso no-alcohólica.
La enfermedad de hígado graso es común, llegando a afectar a un tercio de la población mundial. Sus orígenes son variables, pero están asociados tanto al consumo de alcohol como a la obesidad, la resistencia a la insulina, diabetes mellitus Tipo 2, entre otras. Esteatosis hepática es la primera histopatología observable en la enfermedad, escasamente se desarrolla cirrosis, por lo que la esteatosis persiste, causando hepatomegalia y elevando las concentraciones de las proteínas aminotransferasas alterando el curso de otras enfermedades hepáticas como la hepatitis viral, generando esteatohepatitis, fibrosis, cirrosis y cáncer hepatocelular1.
Estudios previos indican que durante etapas tempranas de la regeneración existe acúmulo graso en hepatocitos2, pero el mecanismo involucrado y cómo podría verse afectado con una esteatosis aún no se comprende.
Para entender mejor los mecanismos y consecuencias de una esteatosis en la regeneración hepática, se plantea la siguiente revisión.
Se ha efectuado una revisión de la literatura médica, para resumir las principales publicaciones relacionadas con la enfermedad de hígado graso no-alcóholica y la regeneración hepática. La selección de artículos relevantes fue guiada por el título y resumen hasta obtener estudios científicos y clínicos originales. La búsqueda de información se realizó en la base de datos MEDLINE (PubMed de marzo a julio del año 2012). La forma de búsqueda se diseñó usando palabras clave identificadas en artículos relevantes según la base de datos consultada en publicaciones disponibles en inglés.
Esteatosis durante la regeneración
Como se ha comprobado en numerosos experimentos, durante los procesos iniciales de la regeneración hepática, resulta normal que los hepatocitos acumulen gotas de grasa en su interior de forma pasajera a las 24 a 72 horas después de una hepatectomía parcial, y tiende a desaparecer por sí misma. Posiblemente esto ocurre como una respuesta metabólica hepatocelular al daño, para generar fuentes de energía y material para la construcción de membranas2; sin embargo, el mecanismo con el que el hígado logra realizarlo permanece aún por dilucidar.
Se ha planteado que la regeneración hepática obedece a un proceso de regeneración celular mediada por cuatro oleadas de replicación: una que exhibe la mayor magnitud, dos de mediana intensidad y una última de baja, las que parecen seguir ritmos circadianos mostrando tener picks mitóticos y que se encuentran relacionadas con tres oleadas de acumulación de grasa hepática, pero aún no se conoce exactamente el mecanismo implicado3.
Múltiples estudios se han realizado para tratar de comprobar el mecanismo de la acumulación de grasas sin mayores resultados, no obstante, recientes estudios han logrado esclarecer que el acúmulo de grasas durante la regeneración hepática no dependería necesariamente de la síntesis de novo en el hígado de ácidos grasos, ni de la reabsorción de estos a nivel intestinal sino que de un transporte desde el tejido adiposo al hígado4.
Rol del tejido graso periférico
Una posibilidad estudiada, es que la esteatosis ocurrida fisiológicamente durante la regeneración hepática es que la grasa provenga del tejido graso periférico. Para ello se evaluaron ratones con lipodistrofias, en estos estudios se muestra nueva evidencia de que las disrupciones en el metabolismo de la glucosa pueden alterar la regeneración hepática, se piensa que la hipoglicemia consecuente a una hepatectomía podría ocasionar una lipólisis en el tejido adiposo que contribuiría al depósito graso en el hígado y, por lo tanto, la regeneración, mostrándose además niveles reducidos de adiponectina de las ratones enfermos, consecuente con antecedentes en los que los niveles de adiponectina están relacionados con la regeneración hepática5.
Características de la enfermedad de hígado graso no-alcohólica (EHGNA)
A pesar de que la enfermedad de hígado graso se conoce desde hace más de 200 años, es recién en 1962 cuando se logra descubrir que existe un tipo de enfermedad de hígado graso que no está relacionada con el alcohol, aunque las características histológicas de ambas enfermedades sean indistinguibles.
La patogenia de la enfermedad de hígado graso no-alcohólica (EHGNA) consiste en dos hits (Figura 1): el primero de ellos es la acumulación de lípidos en los hepatocitos seguido por un segundo hit consistente en la inducción de inflamación, lesión hepatocelular y fibrosis mediado por especies reactivas de oxígeno (ERO), que como se ha visto, tienen un importante papel en los mecanismos de acumulación de grasas de la enfermedad, aunque en este último caso, se habla también de la peroxidación de lípidos, activación del citocromo mitocondrial p450, entre otras múltiples posibles causas1,6,7.
Figura 1. Hits de la patogenia de la enfermedad de hígado graso no-alcohólica (EHGNA)8.
El primer hit, ha sido tema de discusión. Existen múltiples modelos de causas que podrían explicar el acúmulo de grasas en el hígado que van desde modelos hormonales, de estrés oxidativo, mitocondriales hasta modelos adipocéntricos, pero al momento de explicar la acumulación de grasas todos caen en 4 mecanismos generales: la primera es aumentando el ingreso de grasas, la segunda es disminuyendo su salida, la tercera es aumentando su síntesis y la cuarta es disminuyendo su utilización por β-oxidación.
El primer mecanismo, un aumento en la llegada de grasas, podría ocasionar una depleción de los adipocitos, los que serían incapaces de captar metabolitos energéticos circulantes. La insulina resulta ser un importante factor que se encarga de remover los triglicéridos de las lipoproteínas, siendo además un importante factor en el metabolismo graso del hepatocito, posiblemente estos adipocitos depletados ocasionan una resistencia a la insulina para protegerlos de un sobrecargo y evitar el daño tisular y celular. Los adipocitos podrían rebalsar grasa al tejido circundante como músculos e hígado.
Cuando las capacidades de reserva lipídica del hígado, posterior al rebalse de los adipocitos, se encuentra sobreexcedida éste libera ácidos grasos hacia el torrente sanguíneo, acumulándose en órganos y en las paredes de los vasos. Los macrófagos, para tratar de tamponear a los lípidos presentes, los fagocitan y metabolizan, transformándose en células espumosas y liberando, como desechos del metabolismo de los lípidos fagocitados y liberando EROS.
En el segundo caso (un aumento de la ingesta de lípidos), los lípidos deben ser transportados por lipoproteínas desde el tejido adiposo al hígado. Una vez dentro de los hepatocitos, estos requieren una serie de proteínas de transporte intracelular como la FATP o la FAT/CD36, la FABP, las caveolinas-1 entre otras, por lo tanto la acumulación lipídica en los hepatocitos requiere una gran interacción entre múltiples proteínas. Lo que sugeriría que una deficiencia o sobreexpresión en alguna de estas proteínas podría llegar a generar alteraciones en el metabolismo lipídico de los hepatocitos.
En el tercer caso respecto a una disminución de la salida de lípidos, cuando aumenta el ingreso y síntesis de lípidos en el hígado, este a su vez se encarga de aumentar la salida o excreción de estos. El transporte de lípidos depende de la salida de éste a través de lipoproteínas como el VLDL cuya síntesis es regulada a través de los genes de SREBP.
Los receptores biliares de farnesoide X (FXR) regulan la producción y metabolismo tanto de lípidos como de ácidos biliares. Estos últimos regulan la producción de VLDL a través de la proteína triglicérido transferasa (MTTP), posiblemente a través de los mismos FXR. Enfermedades genéticas en el MTTP producen una disminución en la producción de VLDL relacionada con un aumento de placas aterogénicas.
En el cuarto caso, la síntesis en novo de ácidos grasos depende de las enzimas acetil-CoA carboxilasa y de la ácido graso sintasa, de hecho la inhibición de la acetil-CoA carboxilasa podría ser de gran utilidad como terapia para combatir esteatosis hepática.
La regulación de la síntesis de triglicéridos en el hígado depende de la señalización por insulina y la activación del factor de transcripción SREBP-1c que modula la síntesis de lípidos. Un cambio genético que induzca el bypass de la insulina por parte del gen del SREBP-1c podría ocasionar un aumento en los triglicéridos sin aumentar el colesterol. En el caso específico del alcohol, estimula la activación de SREBP a través de señales PPAR. La obesidad y la resistencia a la insulina, al estar relacionadas con procesos inflamatorios y señalización de citoquinas que producen una sobreexpresión de la SREBP. También se encuentra involucrada en la esteatosis hepática la concentración de ácidos biliares, cuyo aumento se traduce en un aumento en la SREBP.
El último caso ocurriría cuando la beta-oxidación se encuentre disminuida y por lo tanto la utilización de los lípidos. La beta-oxidación es un proceso compartimentalizado en las células eucariontes en tres organelos: la mitocondria, el peroxisoma y el retículo endoplasmático, algunos de los procesos enzimáticos claves en estos tres lugares están regulados por la PPAR-α. También se puede asociar esteatosis hepática con un daño mitocondrial, donde el transporte de ácidos grasos se encuentre disminuido6.
El segundo hit se ha asociado a reacciones inmunológicas como las del sistema del complemento, el cual estaría involucrado en la patogenia de múltiples enfermedades como la enfermedad alcohólica de hígado graso y la hepatitis viral. Se ha visto que la activación del complemento está asociada a la EHGNA al daño ocasionado por la activación del complejo ataque membrana (CAM) lo que estaría relacionado con el acúmulo de neutrófilos y el aumento de apoptosis en los hígados enfermos (Figuras 2, 3 y 4)9.
Las concentraciones elevadas de neutrófilos podrían estar asociadas a la secreción de citoquinas proinflamatorias como el TNFα, producido durante respuestas inflamatorias asociadas a la obesidad y que podrían producir el acúmulo de grasas, el TGF β, involucrado en los procesos de fibrosis mediada por las células estrelladas del hígado, la IL-6, que sensibiliza al hígado a la apoptosis, la IL10, cuya deficiencia podría estar relacionada a la progresión de la enfermedad, entre otras citoquinas10.
Otra posible explicación para el segundo hit es una alteración en las adipoquinas, específicamente la visfatina, que es una señal sintetizada por el tejido adiposo blanco que junto a otras señales proinflamatorias como la IL-6, TNF-α y la IL-8 podrían estar relacionadas con la EHGNA. De este modo estudios previos sugieren que la visfatina podría estar co-regulada con la IL-6 a través de feedbacks positivos y negativos11.
Factores de riesgo de la EHGNA
La patogénesis de la EHGNA se encuentra relacionada con la diabetes mellitus tipo 2, efectuar estudios de screaning para esta enfermedad podría servir para realizar una diagnosis precoz e impedir el desarrollo de complicaciones, como enfermedades cardiovasculares. Usualmente se evalúa la presencia de hiperglicemia en la sangre. La sensibilidad de este test depende de la población evaluada. Sin embargo, no se ha establecido cuales pacientes con EHGNA pueden tener una intolerancia a la glucosa oral.
Aproximadamente el 48,7% de los pacientes con esteatosis hepática no alcohólica tuvieron algún tipo de desorden metabólico como la diabetes mellitus Tipo 2. Este grupo tuvo una edad más elevada, mayores índices de masa corporal y menores índices HDL-C12.
Se demuestra que existe una relación entre la desregulación de glucosa y la EHGNA. La edad es un factor importante en el desarrollo de EHGNA junto a elevados índices de masa corporal. Se observa que los pacientes con EHGNA presentan bajos niveles de HDLD-C, niveles séricos de albumina inferiores, lo que puede estar relacionado con un estado avanzado de la enfermedad hepática lo que puede determinar que los desordenes metabólicos, la edad y el índice de masa corporal son importantes factores de riesgo para el desarrollo de una esteatosis hepática no-alcohólica12.
Manifestaciones clínicas de la EHGNA
La esteatosis hepática es un cuadro clínico por lo general asintomático, detectable en el examen físico si es que existe algún tipo de hepatomegalia. Usualmente se puede evaluar al observar un aumento de las aminotransferasas una vez descartada una enfermedad hepática crónica. El radio de valores de la aspartato aminotransferasa y la alanina aminotransferasa es menor en presencia de esteatosis, pero su valor predictivo es pobre en pacientes con una esteatosis severa y fibrosis parenquimal. Los niveles séricos de las fosfatasas alcalinas y la gama-glutamil transferasa están por lo general sobre los niveles normales, estando también elevados los niveles de lípidos séricos y las concentraciones de glucosa en cerca del 75% de los pacientes. Otras posibles anormalidades de laboratorio son la hipoalbuminemia, un tiempo prolongado de protrombina e hiperbilirrubinemia, pero estos son hallazgos extraños en pacientes con una enfermedad avanzada. Otros buenos mecanismos para poder evaluarlo son el radio de cintura y el índice de masa corporal (IMC), que han demostrado ser buenos indicadores para esteatosis hepática7.
A pesar de esto, la esteatosis hepática, en condiciones normales, tienen una prognosis benigna a nivel celular y funcional. Para explicar esto, se plantean distintas teorías: la primera es que de hecho, la esteatosis no produzca daño celular y que los hepatocitos grasos, de hecho, no estén dañados. Otra alternativa es que existan mecanismos compensatorios para reparar el daño hepatocelular.
Posibles señales involucradas en regeneración hepática en EHGNA
La EHGNA severa afecta negativamente la regeneración hepática después de una hepatectomia parcial (Figura 5) lo que no ocurre en un hígado normal o de niveles de esteatosis bajos. En hígados enfermos además se puede apreciar la presencia de altas concentraciones de citoquinas proinflamatorias y una respuesta antioxidante insuficiente que contribuiría a aumentar el daño tisular13.
Existen múltiples teorías que podrían explicarlo. Una de ellas es que la fibrosis producida por el estado avanzado de la EHGNA debido al aumento del estrés oxidativo reduciría las capacidades de los hepatocitos maduros de proliferar, lo que produciría una regeneración hepática dispar8.
Otras teorías explican estos procesos, señalando que la leptina, por ejemplo, es un importante factor en los modelos murinos de esteatosis hepática, ya que, para inducirla, se utilizan o ratones con su gen deletado o knock-out o bien modelos con dietas ricas en grasas.
La deficiencia de leptina en ratones ob/ob produce una regeneración hepática dispar tras una hepatectomía parcial, lo que podría sugerir un papel de la leptina en la falla de la regeneración hepática. No obstante, esto debería reconsiderarse ya que se ha reportado en estudios anteriores un aumento en la tasa de mitosis y una disminución en las señales proapoptóticas y la administración en ratones ob/ob no ha alterado el fenotipo de la deficiencia de leptina y fue incapaz de revertir el déficit en la regeneración de hepatocitos lo que sugiere un papel mucho más complejo de la leptina en los procesos de regeneración7,9.
Un posible factor detonante que podría afectar la regeneración hepática durante procesos de esteatosis debido a su papel en el metabolismo de los lípidos es la ácido graso sintasa. Se ha demostrado que su sobreexpresión se encuentra relacionada con cáncer y en múltiples enfermedades neoplásicas, mientras que su supresión se encuentra relacionada con la disminución en la supervivencia celular, sin embargo, una inhibición de la expresión de ésta, no altera la acumulación de grasas en el hígado durante los períodos de regeneración lo que podría sugerir que su rol resultaría más bien secundario4.
También existe la teoría, que podría explicar una regeneración hepática dispar, a partir de una deficiencia en el retículo endoplásmico de los hepatocitos grasos. El retículo es uno de los organelos celulares encargados de las síntesis, plegamiento de proteínas y del movimiento de calcio intracelular, un aumento en estrés del RE, está asociado a diversas enfermedades crónicas. Estudios en hígados con esteatosis demostraron los RE de los hepatocitos grasos tenían una menor respuesta ante el estrés, sumado al hecho de que la baja expresión de las proteínas chaperonas encargadas del movimiento proteico dentro del organelo y la disminuida expresión de las proteínas encargadas de iniciar señalización proapoptóticas podría explicar la baja tasas de apoptosis en las células grasas15.
Finalmente otra teoría dice que la esteatosis produce algún tipo de modificación en las respuestas inflamatorias necesarias para la regeneración hepática. Un posible candidato es la proteína A20, una inhibidora del factor NfkB, cuya administración en modelos murinos de hepatectomia parcial con esteatosis, demostró corregir los efectos de la regeneración dispar. Posiblemente, esta proteína se encarga de desinhibir la vía de las ciclinas/CDK, a través de la inhibición del factor CDKN1A que es un factor importante para salir de los procesos de diferenciación y proliferación en el hígado. Además demostró también ser un inhibidor potencial de las vías de las leptinas, sin inhibir las caveolinas o los receptores de glucocorticoides, por lo que aporta nuevos indicios de que las vías de las leptinas no están involucradas directamente en los procesos de regeneración16.
Función de las células de Kupffer
Las células de Kupffer son células macrocíticas propias del hígado. Existen diversas teorías que involucran a estas células para explicar la regeneración dispar producida por la esteatosis. Una de ellas implica que la acumulación de ácidos grasos en el interior de las células hepáticas promueve un aumento en las especies reactivas de oxígeno aumentando el estrés oxidativo de la mitocondria y la peroxidación de lípidos. Sumado a esto los hígados con esteatosis poseen una mala eliminación de los radicales libres lo que provocan la acumulación y activación de células de Kupffer, comenzando a producir citoquinas proinflamatorias como el TNF-α y la IL-6 que podrían aumentar el daño hepático (Figura 6).
Las células de Kupffer reconocen patrones asociados a patógenos o bien patrones presentados por células dañadas a través de receptores TLR cuyas proteínas reguladoras (CD14 y TLR4) llevan a una sensibilización frente a los ácidos grasos saturados los que pueden activar la mediada por TLR4 y continuar con la inflamación17,18,19, aunque también reconocen algunas citoquinas como es el caso de la PPARα, que es necesaria para que ocurra la regeneración hepática y está asociado al acúmulo de grasas20.
Estudios previos han demostrado que hígados con una esteatosis leve sin respuesta inflamatoria no presentan diferencia en la regeneración hepática de los hígados sanos, cosa que no ocurrió con hígados con esteatosis severa, viéndose un aumento de las citoquinas proinflamatorias que producen las células de Kupffer ante el daño hepático.
Esto plantea a las células de Kupffer como un posible objeto de estudio en el futuro, explicándose 4 posibles mecanismos en los que estas células pueden estar involucradas en la regeneración y daño hepático por esteatosis:
En este último punto, los ácidos grasos alterados pueden activar a los receptores TLR-4, encargados del reconocimiento de los patrones patológicos exógenos o endógenos de las células de Kupffer y producir respuestas proinflamatorias que desencadenarían y aumentarían el daño hepático.
Aunque no es del todo claro, el colesterol podría tener una relación con la EHGNA debido a que pueden tener algún tipo de daño tisular, se ve que en aumento de LDL se activan células de Kupffer1.
A pesar de que se han realizado múltiples estudios en distintos modelos tanto genéticos cómo bioquímicos, para analizar los efectos de la esteatosis hepática en la regeneración y metabolismo, aún no es posible determinar una vía exacta que altere efectivamente los procesos fisiológicos normales de este órgano.
Se han planteado diversos modelos que relacionan la etiología de la EHGNA con procesos bioquímicos, genéticos o inflamatorios, pero que aún no son posibles de integrar correctamente en un solo gran modelo que la explique y que logre demostrar cómo puede afectar la regeneración, lo que podría resultar muy útil para la práctica clínica y el tratamiento de este síndrome que cada vez es más común en el mundo.
Se plantea la posibilidad de que la esteatosis hepática sea una enfermedad multifactorial en su método de afección en el hígado.
El autor ha completado el formulario de declaración de conflictos de intereses del ICMJE traducido al castellano por Medwave, y declara no haber recibido financiamiento para la realización del artículo/investigación; no tener relaciones financieras con organizaciones que podrían tener intereses en el artículo publicado, en los últimos tres años; y no tener otras relaciones o actividades que podrían influir sobre el artículo publicado. El formulario puede ser solicitado contactando al autor responsable.
El autor señala que la doctora Pamela Romanque U., colaboró en la realización de este artículo.
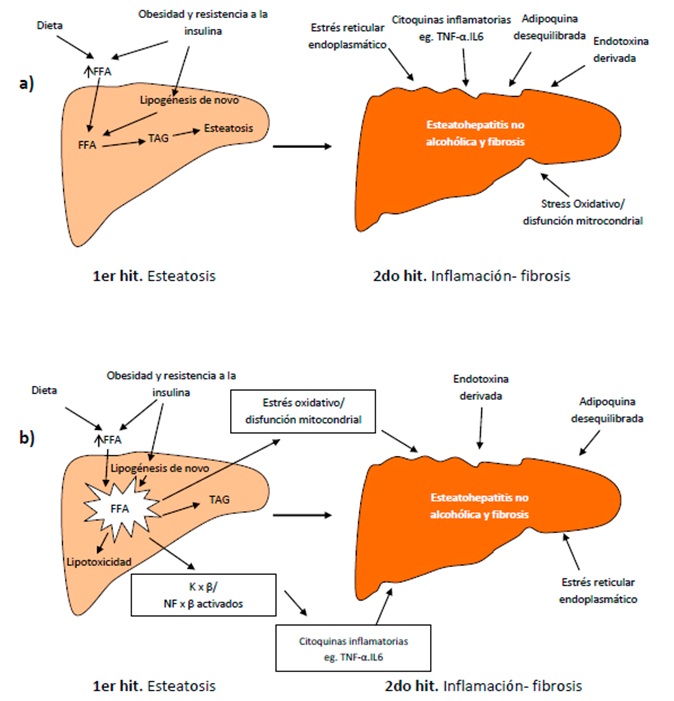 Figura 1. Hits de la patogenia de la enfermedad de hígado graso no-alcohólica (EHGNA)8.
Figura 1. Hits de la patogenia de la enfermedad de hígado graso no-alcohólica (EHGNA)8.
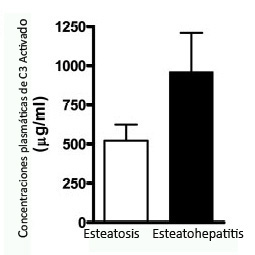 Figura 2. Concentraciones plasmáticas de C3 activado en pacientes con enfermedad de hígado graso no-alcohólica, se puede ver un aumento en las concentraciones de C3 activada en los hígados enfermos9.
Figura 2. Concentraciones plasmáticas de C3 activado en pacientes con enfermedad de hígado graso no-alcohólica, se puede ver un aumento en las concentraciones de C3 activada en los hígados enfermos9.
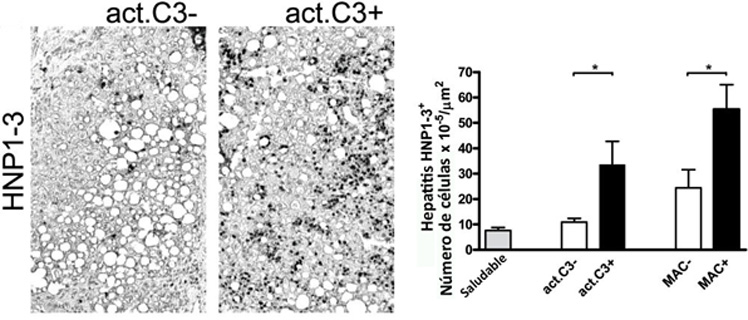 Figura 3. Concentración de HNP1-3 proteína presente en neutrófilos, se puede apreciar una mayor cantidad de neutrófilos en los hígados enfermos, con mayores concentraciones de C3 activada y complejo de ataque membrana9.
Figura 3. Concentración de HNP1-3 proteína presente en neutrófilos, se puede apreciar una mayor cantidad de neutrófilos en los hígados enfermos, con mayores concentraciones de C3 activada y complejo de ataque membrana9.
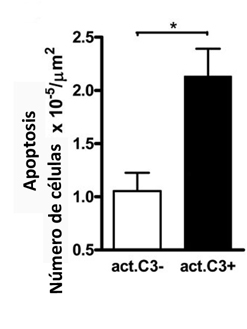 Figura 4. Cantidad de apoptosis en hígados con EHGNA, se puede apreciar un aumento de la apoptosis en hígados enfermos9.
Figura 4. Cantidad de apoptosis en hígados con EHGNA, se puede apreciar un aumento de la apoptosis en hígados enfermos9.
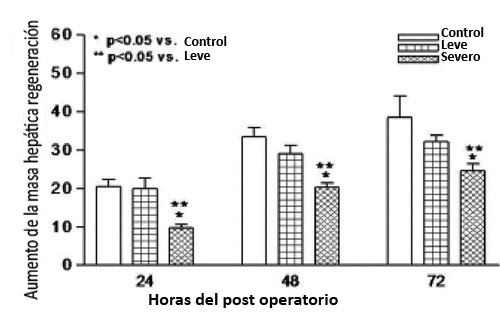 Figura 5. Porcentaje de regeneración hepática a las 24, 48 y 72 horas post hepatectomía parcial de 70% del hígado bajo distintos niveles de esteastosis hepática. Aumenta nivel de esteastosis disminuye la regeneración13.
Figura 5. Porcentaje de regeneración hepática a las 24, 48 y 72 horas post hepatectomía parcial de 70% del hígado bajo distintos niveles de esteastosis hepática. Aumenta nivel de esteastosis disminuye la regeneración13.
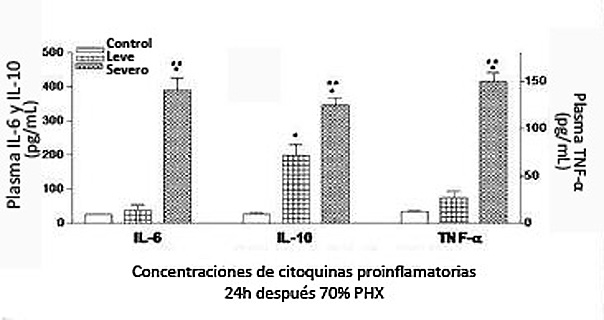 Figura 6. Concentraciones de citoquinas proinflamatorias post hepatectomía parcial en hígados con distintos niveles de esteatosis13.
Figura 6. Concentraciones de citoquinas proinflamatorias post hepatectomía parcial en hígados con distintos niveles de esteatosis13.
 Esta obra de Medwave está bajo una licencia Creative Commons Atribución-NoComercial 3.0 Unported. Esta licencia permite el uso, distribución y reproducción del artículo en cualquier medio, siempre y cuando se otorgue el crédito correspondiente al autor del artículo y al medio en que se publica, en este caso, Medwave.
Esta obra de Medwave está bajo una licencia Creative Commons Atribución-NoComercial 3.0 Unported. Esta licencia permite el uso, distribución y reproducción del artículo en cualquier medio, siempre y cuando se otorgue el crédito correspondiente al autor del artículo y al medio en que se publica, en este caso, Medwave.
Steatosis is the accumulation of fat in hepatocytes, which may be the result of liver regeneration or pathological processes such as alcoholic and nonalcoholic fatty liver disease. Despite its importance, in both cases the exact mechanism that prevails in fatty liver regeneration is poorly understood. Previous studies have shown that patients with fatty liver express dispar regeneration, possibly due to the accumulation of reactive oxygen species generated by inflammatory processes caused by activation of Kupffer cells. In this article we review several factors that affect liver regeneration, trying to understand the underlying mechanism of dispar regeneration in fatty liver as a consequence of nonalcoholic fatty liver disease.
Autor:
Aldo Lagomarsino[1]
Citación: Lagomarsino A. Liver regeneration in nonalcoholic fatty liver disease. Medwave 2012 Dic;12(11):e5559 doi: 10.5867/medwave.2012.11.5559
Fecha de envío: 12/8/2012
Fecha de aceptación: 1/10/2012
Fecha de publicación: 1/12/2012
Origen: no solicitado
Tipo de revisión: con revisión externa por 2 revisores, a doble ciego
Citaciones asociadas
1. Impressum Dic;12(11) Medwave: cuerpo editorial de este número | Link |
Nos complace que usted tenga interés en comentar uno de nuestros artículos. Su comentario será publicado inmediatamente. No obstante, Medwave se reserva el derecho a eliminarlo posteriormente si la dirección editorial considera que su comentario es: ofensivo en algún sentido, irrelevante, trivial, contiene errores de lenguaje, contiene arengas políticas, obedece a fines comerciales, contiene datos de alguna persona en particular, o sugiere cambios en el manejo de pacientes que no hayan sido publicados previamente en alguna revista con revisión por pares.
Aún no hay comentarios en este artículo.
Para comentar debe iniciar sesión
Medwave publica las vistas HTML y descargas PDF por artículo, junto con otras métricas de redes sociales.
Baffy G. Kupffer cells in non-alcoholic fatty liver disease: The emerging view. J Hepatol. 2009 July;51(1): 212–223. | CrossRef | PubMed | PMC |Michalopoulos GK. Liver regeneration after partial hepatectomy critical analysis of mechanistic. Am J Pathol. 2010 Jan;176(1):2-13. Epub 2009 Dec 17. | CrossRef | PubMed | PMC |Zou Y, Bao Q, Kumar S, Hu M, Wang GY, Dai G. Four waves of hepatocyte proliferation linked with three waves of hepatic fat accumulation during partial hepatectomy-induced liver regeneration. PLoS One. 2012;7(2):e30675. Epub 2012 Feb 3.
| CrossRef | PubMed | PMC |Rudnick DA, Davidson NO. Functional relationships between lipidmetabolism and liver regeneration. Int J Hepatol. 2012;2012:549241. | CrossRef | PubMed | PMC |Gazit V, Weymann A, Hartman E, Finck BN, Hruz PW, Tzekov A, et al. Liver Regeneration is Impaired in Lipodystrophic fld Mice. Hepatology. 2010 Dec;52(6):2109-17. | CrossRef | PubMed | PMC |Girija MG. The blind men 'see' the elephant-the many faces of fatty liver disease. World J Gastroenterol. 2008 Feb 14;14(6):831-44. | CrossRef | PubMed | PMC |Veteläinen R, van Vliet A, Gouma DJ, van Gulik TM. Steatosis as a Risk Factor in Liver Surgery. Ann Surg. 2007 Jan;245(1):20-30. | CrossRef | PubMed | PMC |Dowman JK, Tomlinson JW, Newsome PN. Pathogenesis of non-alcoholic fatty liver disease. QJM. 2010 Feb;103(2):71-83. Epub 2009 Nov 13. | CrossRef | PubMed | PMC |Rensen SS, Slaats Y, Driessen A, Peutz-Kootstra CJ, Nijhuis J, Steffensen R, et al Activation of the Complement System in Human Nonalcoholic Fatty Liver Disease. Hepatology. 2009 Dec;50(6):1809-17. | CrossRef | PubMed |Braunersreuther V, Viviani GL, Mach F, Montecucco F. Role of cytokines and chemokines in non-alcoholic fatty liver disease. World J Gastroenterol. 2012 Feb 28;18(8):727-35. | CrossRef | PubMed | PMC |Jarrar MH, Baranova A, Collantes R, Ranard B, Stepanova M, Bennett C, et al. Adipokines and cytokines in non-alcoholic fatty liver disease. Aliment Pharmacol Ther. 2008 Mar 1;27(5):412-21. Epub 2007 Dec 10. | CrossRef | PubMed |Hatziagelaki E, Karageorgopoulos DE, Chounta A, Tsiavou A, Falagas ME, Dimitriadis G. Predictors of impaired glucose regulation in patients with non-alcoholic fatty liver disease. Exp Diabetes Res. 2012;2012:351974. | CrossRef | PubMed | PMC |Veteläinen R, van Vliet AK, van Gulik TM. Severe steatosis increases hepatocellular injury and impairs liver regeneration in a rat model of partial hepatectomy. Ann Surg. 2007 Jan;245(1):44-50. | CrossRef | PubMed | PMC |Newberry EP, Kennedy SM, Xie Y, Luo J, Stanley SE, Semenkovich CF, et al. Altered Hepatic Triglyceride Content After Partial Hepatectomy Without Impaired Liver Regeneration in Multiple Murine Genetic Models. Hepatology. 2008 Oct;48(4):1097-105. | CrossRef | PubMed | PMC |Ben Mosbah I, Alfany-Fernández I, Martel C, Zaouali MA, Bintanel-Morcillo M, Rimola A, et al. Endoplasmic reticulum stress inhibition protects steatotic and non-steatotic livers in partial hepatectomy under ischemia–reperfusion. Cell Death Dis. 2010 Jul 8;1:e52. | CrossRef | PubMed | PMC |Damrauer SM, Studer P, da Silva CG, Longo CR, Ramsey HE, Csizmadia E, et al. A20 Modulates Lipid Metabolism and Energy Production to Promote Liver Regeneration. PLoS One. 2011 Mar 17;6(3):e17715. | CrossRef | PubMed | PMC |Broering R, Lu M, Schlaak JF. Role of Toll-like receptors in liver health and disease. Clin Sci (Lond). 2011 Nov;121(10):415-26. | CrossRef | PubMed |Rivera CA, Adegboyega P, van Rooijen N, Tagalicud A, Allman M, Wallace M. Toll-like receptor-4 signaling and Kupffer cells play pivotal roles in the pathogenesis of non-alcoholic steatohepatitis. J Hepatol. 2007 Oct;47(4):571-9. Epub 2007 Jun 8. | CrossRef | PubMed | PMC |Bertola A, Bonnafous S, Anty R, Patouraux S, Saint-Paul MC, Iannelli A, et al. Hepatic Expression Patterns of Inflammatory and Immune Response Genes Associated with Obesity and NASH in Morbidly Obese Patients. PLoS One. 2010 Oct 22;5(10):e13577. | CrossRef | PubMed | PMC |Matsumoto K, Yu S, Jia Y, Ahmed MR, Viswakarma N, Sarkar J, et al. Critical role for transcription coactivator peroxisome proliferator-activated receptor (PPAR)-binding protein/TRAP220 in liver regeneration and PPARalpha ligand-induced liver tumor development. J Biol Chem. 2007 Jun 8;282(23):17053-60. Epub 2007 Apr 16. | CrossRef | PubMed |
Principales causas y factores asociados a cirrosis hepática en los pacientes del Hospital General de Zona 2 de Chiapas, México 






 Estudios originales
Estudios originales