Revista Biomķdica Revisada Por Pares

 Para Descargar PDF debe Abrir sesi¾n.
Para Descargar PDF debe Abrir sesi¾n.
La matriz ¾sea esta compuesta por dos fases principales a escala nanomķtrica: La orgßnica que representa aproximadamente el 35% del peso seco del hueso. Formada fundamentalmente por proteĒnas, entre las que destaca el colßgeno (90%), tipo I (>95%) y tipo V (<5%). Tambiķn se ha comprobado la presencia, en peque±as proporciones, de colßgeno tipo III, relacionado con las fibras de Sharpey y tipo XII, constituido bajo estrķs mecßnico.
La molķcula de colßgeno contiene a los aminoßcidos hidroxilisina e hidroxiprolina, este ·ltimo un marcador especĒfico de todos los fenotipos de colßgeno (Tabla I). El 10% restante lo forman proteĒnas no colagķnicas (proteoglicanos o glicoproteĒnas).
Tabla 1. ProteĒnas en la matriz ¾sea
En la matriz osteoide hay cuatro tipos de proteoglicanos: hialuronano y condroitĒn sulfato: de molķcula grande, intervienen en las etapas iniciales de la morfogķnesis ¾sea. Biglicano y decorina: de molķcula mßs peque±a, aparecen en las fases siguientes de la formaci¾n ¾sea. Estas proteĒnas no colagķnicas pueden modular la adherencia celular y median en la calcificaci¾n de la matriz orgßnica.
Las proteĒnas con ßcido gamma-carboxi-glutßmico son la osteocalcina y la proteĒna de la matriz con ßcido gamma-carboxiglutßmico. Este ßcido es un aminoßcido que liga calcio y necesita vitamina K para su sĒntesis.
La osteocalcina es una peque±a proteĒna de la matriz sintetizada por los osteoblastos y plaquetas, dependiente de las vitaminas D y K. Representa el 15% de las proteĒnas no colßgenas de la matriz y sus niveles plasmßticos se han considerado como uno de los marcadores bioquĒmicos de la osteogķnesis, relacionßndose con el n·mero y actividad de los osteoblastos.
Las glicoproteĒnas son la osteonectina, la fosfatasa alcalina y las proteĒnas con el tripķptido RGD (Arg-Gly-Asp).
La osteonectina es una glicoproteĒna con gran afinidad por el colßgeno tipo I, por el calcio y por la hidroxiapatita. Representa el 25% de las proteĒnas no colßgenas. Se cree que interviene en la regulaci¾n de la adhesi¾n celular entre la matriz y las cķlulas. En el hueso es necesaria para la mineralizaci¾n normal.
La fosfatasa alcalina es una enzima que libera fosfato inorgßnico a partir de ķsteres fosf¾ricos, necesario para la mineralizaci¾n. Existen varias isoenzimas y, entre ellas la ¾sea, que se han considerado un buen marcador de la actividad osteoblßstica.
ProteĒnas con el tripķptido RGD, tambiķn llamadas SIBLINGS (Small Integrin-Binding Ligand, N-linked Glycoprotein) son fundamentalmente cinco: osteopontina, sialoproteĒnas ¾seas, fibronectina, trombospondina y vitronectina. Son glicoproteĒnas fundamentales en los procesos de remodelado y regeneraci¾n ¾seos, con una secuencia Arg-Gly-Asp (RGD) que es reconocida por las integrinas de los osteoblastos y los osteoclastos (avbeta3, entre otras). Tambiķn act·an como receptores de superficie de las cķlulas ¾seas permitiendo la adhesi¾n de las cķlulas a la matriz extracelular y activando se±ales. Otras proteĒnas procedentes del plasma se encuentran en la matriz orgßnica ¾sea en mayor proporci¾n. Son la alb·mina y la a2-SH-glicoproteĒna, probablemente relacionadas con la incorporaci¾n del calcio a la matriz osteoide.
Por ·ltimo los factores de crecimiento, polipķptidos sintetizados en el propio hueso o procedentes de otros lugares (hĒgado, plaquetas, etc.), que intervienen en la diferenciaci¾n, crecimiento y proliferaci¾n de las cķlulas de forma autocrina o paracrina (1) .
La matriz inorgßnica representa 60 a 70% del peso en seco del hueso. El 99% del calcio, el 85% del f¾sforo, y cerca del 40-60% del sodio y magnesio del organismo residen en el esqueleto. El colßgeno tipo I del osteoide, con una estructura fibrosa tĒpica cuyo dißmetro varia entre 100 y 200 nm es el sustrato en el que, por mineralizaci¾n, se deposita el fosfato cßlcico. Un proceso ordenado, tiempo-dependiente, modulado por las cķlulas, se realiza a raz¾n de 1-2 um al dĒa (2).
La regulaci¾n de esta homeostasis mineral del hueso se lleva a cabo por los iones: calcio, fosfato y magnesio que responden a la acci¾n de la vitamina D3, la hormona paratiroidea (HPT) y la calcitonina. La vitamina D3 aumenta la reabsorci¾n de calcio a nivel intestinal, la HPT la reabsorci¾n a nivel renal, ademßs activa los osteoclastos y la calcitonina, y desactiva los osteoblastos, facilitando la restauraci¾n del nivel basal de calcio.
Los minerales no estßn directamente ligados al colßgeno sino a las proteĒnas no colßgenas. Estas suponen de 3 a 5% de la composici¾n del hueso y proveen emplazamientos activos para la biomineralizaci¾n y uni¾n celular.
Urist (3), en 1965, descubri¾ factores de diferenciaci¾n que facilitaban el reclutamiento y diferenciaci¾n de cķlulas mesenquimales, especĒficamente las que inducen la formaci¾n de osteoblastos, capaces de provocar formaci¾n ¾sea en lugares ect¾picos. Sampath y Reddi (4) observaron que la matriz ¾sea carecĒa de actividad osteoinductora tras la extracci¾n de las proteĒnas que contenĒa, pero que esta actividad era restablecida al reconstituir dicha matriz de nuevo con el extracto proteico, demostrando que las molķculas responsables de la osteoinducci¾n se encontraban en la fracci¾n soluble de la matriz ¾sea. Hecho clave para su posterior aislamiento e identificaci¾n.
A estos factores de diferenciaci¾n se les denomino ProteĒnas Morfogenķticas Humanas (PMH), un grupo de morfogenes que dirigen el desarrollo embriol¾gico de las cķlulas y juegan un papel importante en la fisiologĒa postfetal. Friedenstein (5) describi¾ que pueden dar origen a cinco estirpes celulares distintas: fibroblastos, osteoblastos, condroblastos, adipocitos y mioblastos, en respuesta a diferentes se±ales moleculares que inician la cascada de activaci¾n de genes. Wozney (6) las identific¾ como proteĒnas morfogenķticas ¾seas, parte de la superfamilia de proteĒnas del factor de crecimiento transformador de proteĒnas beta.
Las funciones de las PMH son contradictorias ya que presentan propiedades eclķcticas, una caracterĒstica ·nica de los morfogenes, por lo que se les ha denominado pleiotr¾picas. Los receptores para las PMH son molķculas transmembrana que contienen el dominio serina/treonina/kinasa, de manera que un mismo receptor puede unirse a diferentes PMH (7). La regulaci¾n de sus efectos se debe a que depende de la combinaci¾n de diferentes receptores para desarrollar una acci¾n u otra.
Tienen diferentes orĒgenes. Pueden ser producidos por el sistema inmunitario, por las plaquetas o tambiķn por cķlulas de la lĒnea osteogķnica. Pueden tener su efecto en cķlulas cercanas a las que producen el factor (acci¾n paracrina), sobre las mismas cķlulas (acci¾n autocrina), o pueden ser almacenadas en la matriz ¾sea y ser liberados posteriormente en la resorci¾n ¾sea (acci¾n paracrina tardĒa). Algunas de estas sustancias deben actuar en asociaci¾n con hormonas como la HPT, la calcitonina o la vitamina D3. A continuaci¾n los enumeramos:
Las cķlulas mesenquimßticas pluripotenciales se diferencian directamente en el seno del tejido conjuntivo en cķlulas formadoras ¾seas (osteoblastos) en respuesta a las molķculas de adhesi¾n y los factores solubles de se±al. Este tipo de formaci¾n ¾sea directa es lo que caracteriza el desarrollo del hueso intramembranoso (8).
Las proteĒnas morfogenķticas ¾seas (PMH) promueven la diferenciaci¾n osteoblßstica de las cķlulas pluripotenciales, posiblemente porque exista un gradiente de concentraci¾n de PMH (9). En la secuencia de formaci¾n de hueso endocondral, al que pertenecen la base del crßneo, columna vertebral, pelvis y extremidades, las cķlulas mesenquimßticas siguen un camino hacia la estirpe celular condrogķnica en lugar de hacia la osteogķnica, el motivo tampoco estß bien identificado, pero se especula si las PMH y la sĒntesis local de factores angiogķnicos o angiogenķticos como el Factor bßsico de crecimiento fibroblßstico (FBCF), act·an en esas localizaciones modulando la expresi¾n del fenotipo celular hacia esta lĒnea condrogķnica (10).
Donde se hace mßs evidente este proceso es en los centros de osificaci¾n primaria de los huesos largos, entre la epĒfisis y la metßfisis. En la metßfisis los condrocitos en diferentes estadios de evoluci¾n se disponen ordenadamente en forma de columnas longitudinales, lo que permite el crecimiento del hueso en longitud, a travķs de los centros de osificaci¾n secundaria, cuando los osteoblastos provenientes del pericondrio invaden los espacios dejados por los condrocitos degenerados e inician la formaci¾n ¾sea.
Independientemente de la ruta de la histogķnesis ¾sea, no existe ninguna diferencia bioquĒmica, morfol¾gica o funcional entre el hueso endocondral y el intramembranoso, y durante la reparaci¾n ¾sea del esqueleto en el adulto tambiķn responden a los mismos factores (11). Las diferencias entre estos huesos de distinto origen se encuentran en la carga funcional y la vascularizaci¾n. Por ejemplo, el hueso intramembranoso tiene una vascularizaci¾n mayor que el de las extremidades, lo que puede ser un factor determinante en cuanto a permitir una mayor concentraci¾n de PMH a nivel local (12).
Cuando se produce una fractura ¾sea, las cķlulas y las molķculas de se±al aparecen en el lugar a reparar de la misma manera que en el proceso embriogķnico. En el momento inicial se desencadena una respuesta inflamatoria, con activaci¾n del complemento y rotura de vasos. La degradaci¾n proteolĒtica de la matriz extracelular aporta factores quimiotßcticos para los monocitos y los macr¾fagos.
Una vez que se activan los macr¾fagos liberan el FBCF que estimula a las cķlulas endoteliales a expresar el activador del plasmin¾geno y la procolagenasa. La sangre extravasada forma un coßgulo, y las plaquetas que lo integran tienen una funci¾n dual: la de hemostasia y la de liberar factores como el FCDP y FCT beta. La zona da±ada presenta una hipoxia debido a la rotura de los vasos sanguĒneos. Como consecuencia se produce una disminuci¾n del pH, y los macr¾fagos y leucocitos polimorfonucleares act·an eliminando los detritus celulares a la vez que secretan factores que promueven la quimiotaxis y la mitogķnesis (13).
A los 3-5 dĒas se constituye un tejido de granulaci¾n, consistente en vasos, colßgeno y cķlulas. El colßgeno serß el substrato que contenga los factores a los que serßn sensibles las cķlulas y el lugar donde ellas se anclarßn cuando lleguen a travķs de los vasos, periostio, endostio y mķdula ¾sea, diferencißndose posteriormente en osteoblastos y condroblastos.
La maduraci¾n del tejido de granulaci¾n se produce en varias semanas hasta que se forma el callo ¾seo, mßs tarde sustituido por hueso fibroso inmaduro y posteriormente por hueso lamelar. El papel del callo ¾seo es estabilizar los fragmentos de la fractura ya que si existe movilidad este proceso no puede llevarse a cabo, con lo que el tejido que predominarß serß de tipo cartilaginoso.
El ·ltimo evento que ocurre en la cascada de fen¾menos de reparaci¾n ¾sea es el remodelado. Un proceso de activaci¾n- reabsorci¾n- formaci¾n, donde los osteoclastos se activan produciendo las lagunas de Howship, que serßn repobladas por osteoblastos que expresan osteoide y cuando ķste se calcifica se restaura la morfologĒa ¾sea (14). Este equipo de cķlulas se denomina unidad bßsica multicelular.
El proceso de remodelado en el hueso cortical serß llevado a cabo por los osteoclastos que labran un t·nel que posteriormente se repuebla de osteoblastos. A esta unidad funcional que constituyen ambas cķlulas se le denomina cono de corte y act·a en la matriz como una especie de taladro acompa±ado por las estructuras vasculares que crecen a medida que avanza su actividad erosiva. A cierta distancia del frente de erosi¾n se alinean los osteoblastos, bordeando las paredes de la matriz, que se disponen, en forma progresiva para cerrar el t·nel creado por los osteoblastos pero sin llegar a obliterarlo. El resultado final de todo este proceso serß un conducto de Havers.
El hematoma fracturario y la hipoxia local son los desencadenantes del proceso de consolidaci¾n al que dividiremos en dos fases. La primera caracterizada por la homeostasis y la liberaci¾n de mediadores como el FCDP y FCT beta que promueven la quimiotaxis y la mitogķnesis. En la segunda el colßgeno servirß de sustrato para todos aquellos elementos involucrados en el proceso de cicatrizaci¾n y que llegan a travķs del aporte vascular para diferenciarse en osteoblastos y condroblastos.
Durante esta fase las BMP difunden a partir de esta matriz ¾sea, pasando en una fase posterior a ser sintetizadas por las cķlulas osteogķnicas que han sido reclutadas por las propias BMP y que son sometidas a un proceso de diferenciaci¾n osteoblßstica (Fig. 1). Con ello se consigue el mantenimiento de la acci¾n reparadora sobre el tejido da±ado a lo largo del tiempo necesario para su reparaci¾n (15).
Figura 1. Cascada de formaci¾n del tejido ¾seo.
Las proteĒnas ¾seas morfogenķticas regulan la quimiotaxis, la mitosis, y la diferenciaci¾n. Los factores de transformaci¾n del crecimiento estßn 100 veces mßs concentrados en el hueso que en otros tejidos. Los osteoblastos tienen una concentraci¾n alta de receptores para ķl. Las proteĒnas ¾seas morfogenķticas y los factores de transformaci¾n del crecimiento refuerzan la curaci¾n de la fractura y pueden minimizar la proporciones de no uni¾n (16). Estas proteĒnas tienen una corta vida media biol¾gica y debe mantenerse a concentraciones terapķuticas en el foco de fractura para ser eficaces.
 Tabla 1. ProteĒnas en la matriz ¾sea
Tabla 1. ProteĒnas en la matriz ¾sea
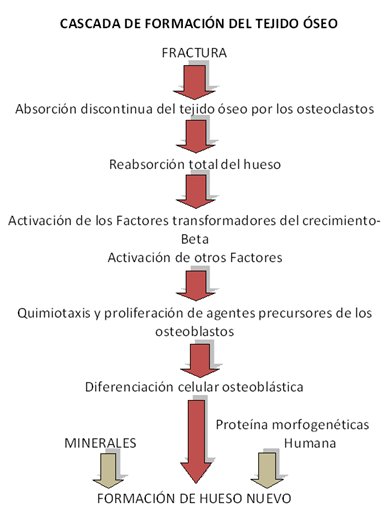 Figura 1. Cascada de formaci¾n del tejido ¾seo.
Figura 1. Cascada de formaci¾n del tejido ¾seo.
 Esta obra de Medwave estß bajo una licencia Creative Commons Atribuci¾n-NoComercial 3.0 Unported. Esta licencia permite el uso, distribuci¾n y reproducci¾n del artĒculo en cualquier medio, siempre y cuando se otorgue el crķdito correspondiente al autor del artĒculo y al medio en que se publica, en este caso, Medwave.
Esta obra de Medwave estß bajo una licencia Creative Commons Atribuci¾n-NoComercial 3.0 Unported. Esta licencia permite el uso, distribuci¾n y reproducci¾n del artĒculo en cualquier medio, siempre y cuando se otorgue el crķdito correspondiente al autor del artĒculo y al medio en que se publica, en este caso, Medwave.
Autor:
Yovanny Ferrer Lozano[1]
Citaci¾n: Ferrer Y. Bone matrix and healing. Medwave 2009 Sep;9(9):4155 doi: 10.5867/medwave.2009.09.4155
Fecha de publicaci¾n: 1/9/2009
Origen: no solicitado
Tipo de revisi¾n: sin revisi¾n por pares
Nos complace que usted tenga interķs en comentar uno de nuestros artĒculos. Su comentario serß publicado inmediatamente. No obstante, Medwave se reserva el derecho a eliminarlo posteriormente si la direcci¾n editorial considera que su comentario es: ofensivo en alg·n sentido, irrelevante, trivial, contiene errores de lenguaje, contiene arengas polĒticas, obedece a fines comerciales, contiene datos de alguna persona en particular, o sugiere cambios en el manejo de pacientes que no hayan sido publicados previamente en alguna revista con revisi¾n por pares.
A·n no hay comentarios en este artĒculo.
Para comentar debe iniciar sesi¾n
Medwave publica las vistas HTML y descargas PDF por artĒculo, junto con otras mķtricas de redes sociales.
Canalis E, Economides AN, Gazzerro E. Bone morphogenetic proteins, their antagonists, and the skeleton. Endocr Rev. 2003 Apr;24(2):218-35. | CrossRef | PubMed |Sim§es R, Carolina C, Verna C, Teixeira R. Cellular and molecular aspects of bone remodelation. Femina 2006; 35(5):279-283. Urist MR. Bone: formation by autoinduction. Science. 1965 Nov 12;150(3698):893-9. | CrossRef | PubMed |Sampath TK, Reddi AH. Dissociative extraction and reconstitution of extracellular matrix components involved in local bone differentiation. Proc Natl Acad Sci U S A. 1981 Dec;78(12):7599-603. | CrossRef | PubMed | PMC |Friedenstein AJ. Precursor cells of mechanocytes. Int Rev Cytol. 1976;47:327-59. | CrossRef | PubMed |Wozney JM, Rosen V, Celeste AJ, Mitsock LM, Whitters MJ, Kriz RW, et al. Novel regulators of bone formation: molecular clones and activities. Science. 1988 Dec 16;242(4885):1528-34. | CrossRef | PubMed |Nunes FD, Valenzuela Mda G, Rodini CO, Massironi SM, Ko GM. Localization of Bmp-4, Shh and Wnt-5a transcripts during early mice tooth development by in situ hybridization. Braz Oral Res. 2007 Apr-Jun;21(2):127-33. | CrossRef | PubMed |De Biase P, Capanna R. Clinical applications of BMPs. Injury. 2005 Nov;36 Suppl 3:S43-6. | CrossRef | PubMed |Ferrigno C, Della N, Fantoni D. A comparative study of osteosynthesis with plates and plates associated with grafts of bone morphogenic proteins (Gen-Tech«) in distal radio-ulna fractures in dogs with less than 6 quilograms. Pesq. Braz j vet res 2007 ;27(2):65-69. Hutmacher DW, Garcia AJ. Scaffold-based bone engineering by using genetically modified cells. Gene. 2005 Feb 28;347(1):1-10. | CrossRef | PubMed |Murugan R, Ramakrishna A. Development of nanocomposites for bone grafting. Composites Science and technology 2005; 65: 2385-406. | CrossRef |Khan SN, Cammisa FP Jr, Sandhu HS, Diwan AD, Girardi FP, Lane JM. The biology of bone grafting. J Am Acad Orthop Surg. 2005 Jan-Feb;13(1):77-86. | PubMed |Selvamurugan N, Jefcoat SC, Kwok S, Kowalewski R, Tamasi JA, Partridge NC. Overexpression of Runx2 directed by the matrix metalloproteinase-13 promoter containing the AP-1 and Runx/RD/Cbfa sites alters bone remodeling in vivo. J Cell Biochem. 2006 Oct 1;99(2):545-57. | CrossRef | PubMed |Gil J, Garrido R, Gil R, Melgosa M. Materiales para la reparaci¾n y sustituci¾n ¾sea. Factores de crecimiento y terapia genķtica en CirugĒa Ortopķdica y TraumatologĒa. Mapfre Medicina 2003;14: 51-65. | Link |Bostrom MP. Expression of bone morphogenetic proteins in fracture healing. Clin Orthop Relat Res. 1998 Oct;(355 Suppl):S116-23. | CrossRef | PubMed |Zambuzzi W, Oliveira RC, Piozzi R, Cestari T, Taga R. Histological evaluation of acellular and demineralized fetal bovine bone in the subcutaneous of rats. Rev Bra ortop 2006;41(6): 227-232. 






 Estudios originales
Estudios originales