Revista Biomķdica Revisada Por Pares

 Para Descargar PDF debe Abrir sesi¾n.
Para Descargar PDF debe Abrir sesi¾n.
Este texto completo es la transcripci¾n editada y revisada de una conferencia dictada en reuni¾n clĒnica del Departamento de Medicina, Hospital ClĒnico Universidad de Chile. El director del Departamento de Medicina es el Dr. Alejandro Cotera y la coordinadora de las reuniones clĒnicas es la Dra. Miriam Alvo.
El hierro es un elemento muy abundante en la naturaleza, sin embargo el estado en que se encuentra hace que para los organismos vivos sea muy difĒcil obtenerlo; por esta raz¾n los individuos estßn provistos de eficaces mecanismos para ahorrar el hierro que poseen al nacer.
El hierro es un elemento esencial para la vida: participa en procesos vitales como la sĒntesis del grupo hem, constituyente de la molķcula de hemoglobina que se encarga del transporte de oxĒgeno por el organismo; ademßs interviene en todas las reacciones con transferencias de electrones, especialmente la fosforilaci¾n oxidativa y contribuye en otros procesos de biosĒntesis, incluyendo el de los ßcidos nucleicos. No obstante, el hierro tambiķn es potencialmente muy t¾xico, por lo que nunca se encuentra en su estado de ion libre circulando por el organismo: a travķs de todo el proceso de absorci¾n, utilizaci¾n y almacenamiento el hierro siempre se encuentra ligado a proteĒnas, especialmente en su forma ferrosa; de lo contrario podrĒa desencadenar la producci¾n de radicales super¾xidos muy da±inos, tanto para las proteĒnas como para los ßcidos nucleicos. El hierro circula unido a su transportador, la transferrina; entra a la cķlula a travķs de los receptores de transferrina; sale desde los endosomas lisosomales en un proceso mediado por la proteĒna DMT1, que es una proteĒna transportadora de metales bivalentes; se almacena por medio de la ferritina y finalmente sale de las cķlulas gracias a la mediaci¾n de otra proteĒna de membrana, la ferroportina, que es una exportadora de membrana tambiķn llamada IREG 1 u OMTP. Todo el proceso estß conducido por alguna proteĒna, sea transportadora o de dep¾sito.
Todas las proteĒnas que participan en el metabolismo del hierro estßn reguladas a su vez por los niveles de este elemento, tanto a nivel de la inducci¾n de los genes que las codifican como en su regulaci¾n post-transcripcional, es decir, cuando ya se ha sintetizado el ßcido ribonucleico mensajero (ARNm). Esto es especialmente vßlido para la sĒntesis de la ferritina, que es la proteĒna de dep¾sito y para la molķcula del receptor de transferrina, que tienen una regulaci¾n antag¾nica entre ellas, de modo que cuando el organismo dispone de abundante cantidad de hierro aumenta la sĒntesis de ferritina y, por el contrario, si disminuye la cantidad de hierro aumenta la sĒntesis del receptor de transferrina. Esta regulaci¾n se realiza a nivel del ARNm de ambas proteĒnas por acci¾n de las molķculas ōrespondedoras al hierroö, que act·an como sensores del hierro intracelular. Este es un mecanismo de regulaci¾n recĒproca muy complejo e interesante. Las otras proteĒnas que participan en el transporte intracelular del hierro y que son responsables de la absorci¾n, salida del endosoma (proteĒna DMT1) y salida celular mediada por ferroportina tambiķn tienen una regulaci¾n dependiente de hierro, pero por otros mecanismos.
El primer paso del metabolismo del hierro es la absorci¾n en el intestino, especĒficamente en el duodeno, a nivel de los enterocitos ubicados en el ßpice de las vellosidades, llamados enterocitos maduros. En los enterocitos de la cripta se ubicarĒan los censores de niveles de hierro en el organismo, cuya informaci¾n llega de alguna manera al enterocito para regular la absorci¾n.
El hierro que ingresa al organismo es tipo no hem, inorgßnico y se debe reducir desde su estado normalmente fķrrico, trivalente, hacia su estado ferroso para ser absorbido; en este proceso intervienen dos factores: el primero es la acidez gßstrica, y aunque se sabe que la aquilia gßstrica conduce a anemia ferropriva, se suele olvidar este concepto que ha adquirido relevancia nuevamente en el ·ltimo tiempo; el otro factor que interviene en la reducci¾n del hierro fķrrico a ferroso es la proteĒna de membrana Dcytb, una enzima reductasa que pertenece a la familia de citocromos b56 cuya sĒntesis es fuertemente inducida por la deficiencia de hierro, al igual que la proteĒna DMT1 transportadora de cationes y se postula que serĒa la ·nica capaz de absorber el hierro desde el intestino. Actualmente se sabe que la mutaci¾n homocig¾tica de DMT1 induce un cuadro de anemia microcĒtica hipocroma congķnita severa y que se asocia a sobrecarga de hierro en los macr¾fagos, porque DMT1 tambiķn es muy importante en el reciclamiento del hierro a travķs de ķstos. Finalmente, como esta cķlula recibe se±ales de la cantidad de hierro que existe en el individuo, el hierro que sale de la cķlula a travķs de la ferroportina se almacena en la ferritina, lo que constituye un mecanismo de regulaci¾n, ya que ese hierro unido a la ferritina se perderß por las deposiciones con la descamaci¾n intestinal normal.
El hierro no sale de la cķlula en forma libre, sino unido a la proteĒna exportadora de membrana ferroportina, que cumple un rol fundamental. Una vez que el hierro sale de la cķlula nuevamente se debe convertir desde hierro ferroso, que es el estado intracelular, hacia hierro fķrrico, que es menos perjudicial que el ferroso; ademßs la transferrina, proteĒna transportadora plasmßtica del hierro, s¾lo acepta al hierro en su estado fķrrico. El proceso lo realiza la proteĒna haphaestin, que comparte 50% de homologĒa con la ceruloplasmina, ya que cumple la misma funci¾n en el macr¾fago cuando el hierro debe salir de ķste.
El cĒrculo del metabolismo del hierro se completa con el reciclamiento de este elemento gracias a la fagocitosis de los eritrocitos senescentes, proceso que se lleva a cabo bajo la acci¾n de una serie de enzimas y cambios de PH que ocurren en el interior del fagosoma, el que finalmente libera el hierro, entre otras molķculas, a partir de otra enzima que es la hem-oxigenasa. Este proceso protagonizado por el macr¾fago libera alrededor de 20 a 25 miligramos diarios de hierro elemental y sirve de sustrato para que la mķdula ¾sea produzca 300 billones de eritrocitos diarios en el ser humano normal; el mecanismo de absorci¾n intestinal tiene como funci¾n reponer las pķrdidas diarias de hierro, que ascienden a 1 a 2 mg aproximadamente, mientras que el hierro que se utiliza en la producci¾n de los eritrocitos es el que se estß reciclando. Por eso se dice que organismo humano es muy ahorrador de hierro.
El equilibrio entre absorci¾n intestinal y reciclamiento es el ·nico mecanismo de regulaci¾n de los niveles de hierro en el organismo, ya que no existe un mecanismo de excreci¾n; s¾lo la sincronizaci¾n muy fina entre ambos procesos permite mantener la cantidad de hierro estable dentro del cuerpo. Actualmente se sabe que la hormona que regula esta coordinaci¾n entre el enterocito y el macr¾fago es la hepcidina, cuyo nombre se compone de hep, porque se sintetiza a nivel hepßtico, y cidina, porque se descubri¾ cuando se estaban estudiando factores con propiedades antimicrobianas de la inmunidad innata.
La hepcidina, molķcula peptĒdica de 25 aminoßcidos, regula la mantenci¾n de la cantidad de hierro del organismo; para ejercer este efecto se une a la ferroportina e induce su internalizaci¾n y posterior degradaci¾n, es decir, la hepcidina bloquea la entrega de hierro desde el enterocito hacia el plasma y tambiķn desde el macr¾fago hacia el plasma, lo que se traduce en menor disponibilidad de hierro para ser utilizado por el organismo. En resumen, la hepcidina es la proteĒna que regula la disponibilidad de hierro en el individuo: cuando aumenta su nivel disminuye el hierro disponible y cuando su nivel disminuye, la cantidad de hierro disponible aumenta.
La expresi¾n del gen de la hepcidina es controlada tanto por los niveles de hierro como por la inflamaci¾n, que permiten ajustar la cantidad de hierro circulante seg·n los requerimientos. Con respecto a la inflamaci¾n, las citoquinas inflamatorias probablemente convergen en una vĒa final com·n con la interleuquina 6, que desencadena una cascada de se±ales que permiten que el gen de la hepcidina se exprese. Por otra parte, la sobre expresi¾n de la hepcidina explica el patr¾n que se observa en la inflamaci¾n de hiposideremia, es decir, niveles de hierro circulante disminuidos, con ferritina aumentada como expresi¾n de los dep¾sitos aumentados en los macr¾fagos; lo anterior se explica por la degradaci¾n de la ferroportina, lo que impide la salida del hierro desde los enterocitos y macr¾fagos. En resumen, hipoferremia y ferritina aumentada es el patr¾n tĒpico de la anemia inflamatoria. Aparte de lo se±alado existen otros factores que regulan la sĒntesis de hepcidina: disminuyen su producci¾n la anemia, la hipoxia y la actividad eritropoyķtica. Un fen¾meno interesante es que los pacientes talasķmicos o con sĒndrome mielodisplßsico que tienen hierro en exceso tienen niveles de hepcidina inadecuadamente bajos; esto se explica porque la sĒntesis de esta ·ltima es inhibida por la actividad eritropoyķtica, lo que provoca una sobrecarga de hierro a pesar de que ķste se encuentre en niveles excesivos, es decir, la absorci¾n de hierro contin·a debido a que los niveles de hepcidina se encuentran inadecuadamente bajos. Esto demuestra que el estĒmulo eritropoyķtico es un factor inhibidor de la sĒntesis de hepcidina mucho mßs potente que el estĒmulo de sobrecarga de hierro, por mecanismos que a·n se desconocen.
Mßs interesante a·n es el hecho de que la hepcidina es el factor com·n de todas las variedades de hemocromatosis: las cuatro variantes de hemocromatosis recesiva y la ·nica presentaci¾n dominante, que implica la mutaci¾n de la ferroportina, tienen en com·n bajos niveles de hepcidina, debido a que todas las proteĒnas que mutan en la hemocromatosis participan en la regulaci¾n que el hierro ejerce sobre la expresi¾n del gen de hepcidina a nivel de la membrana del hepatocito y la mayorĒa de estas proteĒnas funcionan como co-receptores, en un mecanismo de se±alizaci¾n muy complejo. Cuanto mßs bajos son los niveles de hepcidina, mßs grave es la sobrecarga de hierro. Por otra parte, las cuatro principales proteĒnas mutadas en la forma clßsica de esta patologĒa, dentro de las cuales destaca la HFE, pero tambiķn las del gen de la hepcidina, del receptor II de la transferrina y la hemosiderina, participan en la regulaci¾n de la sĒntesis de hepcidina por el hierro. Desde un punto de vista filogenķtico es mßs problemßtica la sobrecarga de hierro que la deficiencia de ķste, como lo demuestran los estudios efectuados en ratas expuestas a infusiones de hepcidina, en las cuales disminuye en forma importante la sobrecarga de hierro, lo que sugiere que esta terapia serĒa mucho mßs eficaz que utilizar quelantes de hierro en estas condiciones clĒnicas.
En la figura 1 se observa el proceso completo del metabolismo del hierro. En el paso 1, el hierro que se requiere para la eritropoyesis, en cantidad de 20 a 25 mg/dĒa, es provisto por la destrucci¾n de los gl¾bulos rojos senescentes a travķs de los macr¾fagos. En la etapa 2, el hierro liberado al plasma por la ferroportina es oxidado por la ceruloplasmina y transportado por la transferrina a los precursores eritroides de la mķdula ¾sea. En el paso 3, el hierro es absorbido a nivel intestinal por los enterocitos del duodeno, lo que compensa las pķrdidas diarias de 1 a 2 mg/dĒa. La hepcidina, pķptido rico en cisteĒna sintetizado por los hepatocitos y representado en la etapa 4, regula negativamente la exportaci¾n de hierro desde los enterocitos (fase 5) y desde los macr¾fagos a travķs de la uni¾n a ferroportina (fase 6), induciendo la internalizaci¾n y degradaci¾n.
Figura 1. Metabolismo del hierro.
La anemia ferropriva y la de las enfermedades cr¾nicas, o anemia inflamatoria, son las anemias que se observan con mayor frecuencia en la prßctica clĒnica. La deficiencia de fierro es muy com·n: la OMS se±ala que hasta 23% de las mujeres embarazadas tienen deficiencia de hierro, lo cual no significa que estķn anķmicas; ademßs se encuentra este problema en 4% de los hombres entre 15 y 59 a±os, en 10% de las mujeres entre 15 y 59 a±os y en 12% de los mayores de 60 a±os. En la actualidad es muy raro observar las caracterĒsticas clĒnicas clßsicas de la deficiencia grave de hierro, como glositis, queilitis, disfagia y coiloniquia; el reconocimiento clĒnico de la deficiencia de hierro hoy en dĒa se basa en determinaciones de laboratorio. En la poblaci¾n adulta, las mujeres en edad reproductiva son las mßs susceptibles, ya que la pķrdida menstrual puede llegar a 42 mg de hierro por ciclo y un embarazo consume 700 mg.
Es importante recordar que la deficiencia de hierro ocurre cuando se produce un desbalance entre las necesidades y el suministro. Esto puede ocurrir, en primer lugar, debido a demandas no cubiertas por el hierro absorbido, como ocurre cuando hay aumento de los requerimientos, dietas demasiado estrictas y en presencia de sĒndromes de malaabsorci¾n; por ejemplo, puede haber aumento de requerimientos en ni±os, mujeres embarazadas, adolescentes en crecimiento rßpido o personas bajo entrenamiento muscular intenso. El desbalance entre necesidades y suministro se puede deber, en segundo lugar, a aumento de las pķrdidas de hierro, sea fisiol¾gicas, como las que ocurren por la menstruaci¾n y el embarazo, o por pķrdidas gastrointestinales que se manifiestan como pķrdida de sangre oculta en las deposiciones, que explicarĒa 62% de los casos de anemia en la poblaci¾n adulta. Tambiķn puede haber pķrdidas de hierro por otras vĒas, no gastrointestinales, pero son menos frecuentes. Por lo tanto, el diagn¾stico de anemia ferropriva no es definitivo sino que es el punto de inicio de un estudio que debe continuar hasta que se identifique la causa.
La anemia ferropriva es el estado final de la deficiencia de hierro, por lo tanto cuando se trata al paciente se corrige primero la anemia, luego se recuperan los niveles de hemoglobina y finalmente se rellenan los dep¾sitos. Desde el punto de vista clĒnico, hay dos etapas importantes de reconocer en este proceso: una es la depleci¾n de los dep¾sitos, en la cual el paciente no tiene anemia y se evidencia por la disminuci¾n de la ferritina sķrica, que es el parßmetro que tiene mejor correlaci¾n con los dep¾sitos de hierro; ademßs, se considera que un valor de ferritina menor de 12 mcg/L es muy especĒfico de deficiencia de hierro, pues s¾lo existen dos situaciones en las cuales este valor se encuentra bajo sin alteraci¾n de los dep¾sitos: el hipotiroidismo, que es muy fßcil de reconocer clĒnicamente y la deficiencia de ßcido asc¾rbico, que es muy poco frecuente. Sin embargo todos los autores recomiendan emplear un valor de corte mßs alto en la prßctica clĒnica: algunos sugieren 30 mcg/L y otros, 40 mcg/L, para aumentar la sensibilidad. La gran limitaci¾n de este parßmetro es que se trata de un reactante de fase aguda, por lo tanto aumenta en forma inespecĒfica cuando hay inflamaci¾n y pierde su valor como reflejo de los dep¾sitos.
La siguiente etapa en el proceso de pķrdida de hierro es lo que se conoce como eritropoyesis deficiente en hierro, en la cual no s¾lo disminuye la ferritina, sino tambiķn la sideremia o ferremia. Habitualmente esto se acompa±a de aumento compensatorio de los niveles de transferrina, que es la proteĒna transportadora, y de disminuci¾n de la saturaci¾n de transferrina; este patr¾n no s¾lo refleja una deficiencia de los dep¾sitos, sino tambiķn una repercusi¾n sobre la producci¾n de gl¾bulos rojos, pero habitualmente el paciente a·n no tiene anemia; cuando ķsta aparece, por supuesto que ya existen las dos condiciones se±aladas previamente. Sin embargo, la ferremia y la transferrina no s¾lo se alteran por la inflamaci¾n, sino tambiķn por otras condiciones, lo que determina que su rendimiento diagn¾stico, sensibilidad y especificidad sean relativamente bajos; por ello se considera que el examen mßs confiable para evaluar el estado de eritropoyesis deficiente en hierro, es la cuantificaci¾n de los niveles del receptor soluble de la transferrina, que no se modifica frente a la inflamaci¾n y aumenta muy precozmente cuando el hierro disponible para la sĒntesis de gl¾bulos rojos disminuye.
El diagn¾stico de anemia ferropriva se establece en presencia de anemia microcĒtica e hipocroma, es decir, con volumen corpuscular medio disminuido, habitualmente menor de 80 um3 y hemoglobina corpuscular media disminuida, que indica menor carga de hemoglobina en los gl¾bulos rojos. Un parßmetro que se cita con frecuencia, pero se utiliza poco es el ancho de distribuci¾n eritrocitario (ADE), que en la talasemia, la otra gran causa de anemia microcĒtica e hipocroma, estß disminuido. En la anemia ferropriva el perfil de hierro se caracteriza por ferremia disminuida, capacidad de fijaci¾n de hierro aumentada y ferritina disminuida. La muestra se debe tomar siempre en ayunas por la ma±ana, ya que hay un ritmo circadiano para la ferremia. Finalmente, lo mßs importante es el diagn¾stico de la causa.
Dentro del diagn¾stico diferencial de la anemia ferropriva se debe considerar a las anemias microcĒticas hipocr¾micas no hiposiderķmicas, fundamentalmente las talasemias, que no son frecuentes en Chile pero sĒ en otros paĒses. Para los pacientes talasķmicos el aporte de hierro es perjudicial, ya que tienen una sobrecarga basal, por lo tanto es importante tener en mente esta posibilidad y ante cualquier duda confirmar que la anemia es por deficiencia de hierro antes de iniciar el tratamiento.
El gran problema se presenta frente a la anemia de la enfermedad cr¾nica, que se caracteriza por hiposideremia y cuando es de larga evoluci¾n tambiķn es microcĒtica e hipocr¾mica, debido a una deficiencia de hierro de tipo funcional; por lo tanto es difĒcil hacer el diagn¾stico diferencial, especialmente cuando el paciente que tiene anemia por enfermedad cr¾nica tiene ademßs una deficiencia de hierro sobreagregada: en general se puede diferenciar una anemia pura por deficiencia de hierro de una anemia inflamatoria, pero el problema es cuando ambas coexisten. En estos enfermos se recomienda determinar el cociente entre el receptor soluble de la transferrina y el logaritmo de la ferritina; si el resultado es menor que uno es una anemia inflamatoria pura y si es mayor que dos significa que ademßs hay una deficiencia de hierro. Muchas condiciones inflamatorias cr¾nicas se benefician al corregir la deficiencia de hierro asociada.
De los exßmenes se±alados previamente, el que tiene mayor rendimiento es la ferritina: valores menores de 15 mcg/L se asocian a deficiencia de hierro con una probabilidad muy superior a cualquiera de las otras determinaciones, como volumen corpuscular medio, hemoglobina, hemoglobina corpuscular media y saturaci¾n de transferrina, tanto en poblaci¾n general como en ancianos (2).
En la segunda parte de esta conferencia se hablarß sobre el diagn¾stico etiol¾gico de la anemia ferropriva y su tratamiento.
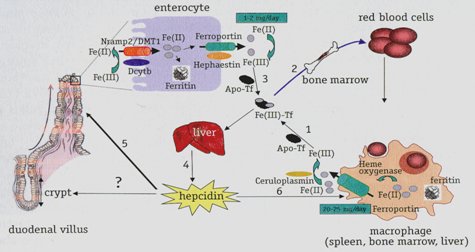 Figura 1. Metabolismo del hierro.
Figura 1. Metabolismo del hierro.
 Esta obra de Medwave estß bajo una licencia Creative Commons Atribuci¾n-NoComercial 3.0 Unported. Esta licencia permite el uso, distribuci¾n y reproducci¾n del artĒculo en cualquier medio, siempre y cuando se otorgue el crķdito correspondiente al autor del artĒculo y al medio en que se publica, en este caso, Medwave.
Esta obra de Medwave estß bajo una licencia Creative Commons Atribuci¾n-NoComercial 3.0 Unported. Esta licencia permite el uso, distribuci¾n y reproducci¾n del artĒculo en cualquier medio, siempre y cuando se otorgue el crķdito correspondiente al autor del artĒculo y al medio en que se publica, en este caso, Medwave.
Este texto completo es la transcripci¾n editada y revisada de una conferencia dictada en reuni¾n clĒnica del Departamento de Medicina, Hospital ClĒnico Universidad de Chile. El director del Departamento de Medicina es el Dr. Alejandro Cotera y la coordinadora de las reuniones clĒnicas es la Dra. Miriam Alvo.
Author:
Daniel Araos H.[1]
Citaci¾n: Araos D. Iron deficiency anemia I: iron metabolism, diagnosis of iron deficiency anemia. Medwave 2009 Sep;9(9):e4154 doi: 10.5867/medwave.2009.09.4154
Fecha de publicaci¾n: 1/9/2009
Nos complace que usted tenga interķs en comentar uno de nuestros artĒculos. Su comentario serß publicado inmediatamente. No obstante, Medwave se reserva el derecho a eliminarlo posteriormente si la direcci¾n editorial considera que su comentario es: ofensivo en alg·n sentido, irrelevante, trivial, contiene errores de lenguaje, contiene arengas polĒticas, obedece a fines comerciales, contiene datos de alguna persona en particular, o sugiere cambios en el manejo de pacientes que no hayan sido publicados previamente en alguna revista con revisi¾n por pares.
A·n no hay comentarios en este artĒculo.
Para comentar debe iniciar sesi¾n
Medwave publica las vistas HTML y descargas PDF por artĒculo, junto con otras mķtricas de redes sociales.







 Estudios originales
Estudios originales