Revista Biomédica Revisada Por Pares

Este texto completo es la transcripción editada y revisada de una conferencia dictada en el Curso Desafíos y Oportunidades en Gastroenterología y Nutrición, organizado por la Sociedad Chilena de Pediatría durante los días 3, 4 y 5 de mayo de 2007. Directores: Dra. Sylvia Cruchet y Dr. Francisco Moraga.
Los últimos adelantos en manometría, cintigrafía e imágenes han ayudado mucho al estudio de los trastornos de la motilidad digestiva, que afectan a más de 15% de la población a nivel mundial. En esta presentación se revisarán los aspectos anatómicos y fisiológicos del sistema digestivo más relevantes para comprender y manejar las condiciones derivadas de las alteraciones de la motilidad digestiva.
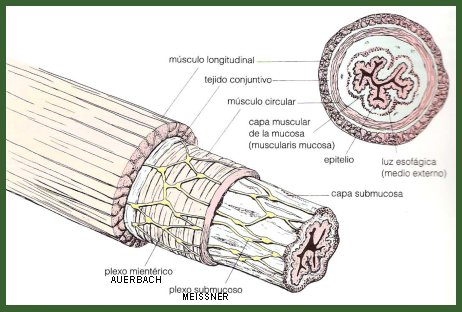
Figura 1. Estructura de la pared del tubo digestivo.
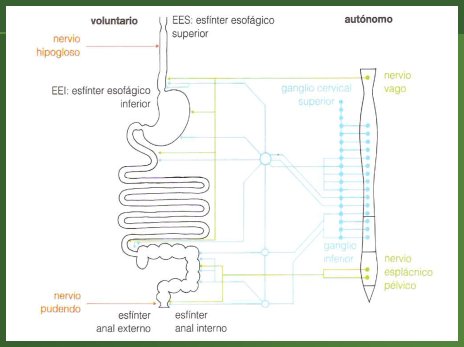
La inervación tiene un componente extrínseco, que a su vez tiene los subcomponentes voluntario y autónomo. El componente voluntario depende del nervio hipogloso, que regula el acto de la deglución y del nervio pudendo, que se relaciona con el manejo del esfínter externo, formado por músculo estriado (Fig. 2).
Figura 2. Inervación extrínseca del tubo digestivo: componentes voluntario y autónomo
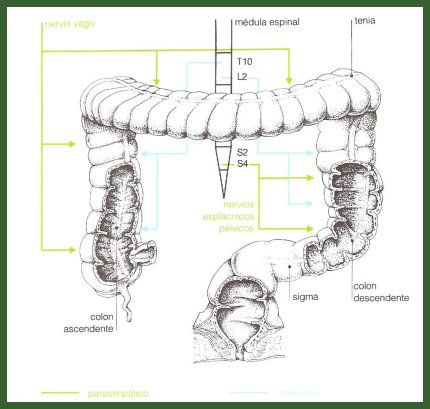
El componente autónomo está dado por el sistema nervioso simpático y parasimpático. El parasimpático inerva y estimula el tubo digestivo, tanto en la parte motora como en su función secretora, hasta alrededor de la mitad del colon transverso, a través del nervio vago, que nace en el encéfalo; la otra mitad es inervada por el sistema simpático a través del nervio pélvico, que proviene de la médula espinal (Fig. 3).
Figura 3. Inervación extrínseca del tubo digestivo: componente autónomo.
El sistema nervioso simpático funciona con base en una primera neurona, que se ubica en la médula y una segunda neurona, que se ubica en los ganglios paravertebrales; de éstos salen fibras nerviosas eferentes hacia todo el tubo digestivo y llegan a las terminaciones, ya sea del sistema nervioso entérico o cerebro intestinal, o directamente a las fibras musculares. El sistema nervioso simpático es fundamentalmente inhibitorio y su principal neurotransmisor es la norepinefrina, así como el principal neurotransmisor del sistema nervioso parasimpático es la acetilcolina.
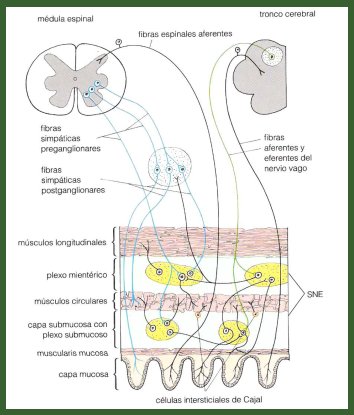
El sistema nervioso intrínseco tiene tres componentes: las células intersticiales de Cajal, el plexo mientérico de Meissner, ambos situados en la submucosa y el plexo de Auerbach, situado entre las dos capas musculares; el conjunto constituye el denominado cerebro intestinal, que se compone de ocho a diez millones de neuronas, con sus correspondientes neurotransmisores, que se comunican con el encéfalo a través del nervio vago, que entrega información tanto a los plexos como a los músculos, mediante un sistema de vías eferentes, que permiten efectuar la contracción, y un sistema de vías aferentes, que llevan las sensaciones y la percepción del dolor al encéfalo (Fig. 4).
Figura 4. Inervación intrínseca del tubo digestivo.
Las células de Cajal se ubican en la parte más profunda de la submucosa, en íntima relación con las fibras musculares circulares y tendrían una función de marcapaso del ritmo intestinal, que en el estómago es de 3 oscilaciones por minuto y en el intestino, entre 8 y 10. El número de células de Cajal varía a lo largo del tubo digestivo: aumenta de proximal a distal, de modo que la mayor cantidad se encuentra en el intestino delgado, es decir, donde las contracciones peristálticas son más intensas y más frecuentes hay mayor cantidad de células de Cajal.
El sistema nervioso entérico autónomo, o cerebro intestinal, cumple su función mediante neurotransmisores, que en este caso se llaman mensajeros, que facilitan la transmisión de información desde el sistema nervioso parasimpático y simpático a las células ganglionares o, directamente, a las fibras musculares. Estos mensajeros se clasifican en estimuladores e inhibidores; los estimuladores se relacionan con el parasimpático y entre ellos se cuentan neurotransmisores clásicos, como la serotonina (5-HT) y algunos péptidos y hormonas intestinales, entre ellas la motilina. Muchos de los medicamentos que se utilizan en la actualidad actúan sobre estos neurotransmisores; por ejemplo, entre los agonistas de la motilina está la eritromicina, que se utiliza para estimular el vaciamiento gástrico y mejorar las paresias gástricas; también hay agonistas de la serotonina, como el tegaserod, que se usa en colon irritable con estreńimiento. Por otra parte, los mensajeros inhibidores se relacionan con el sistema simpático; entre ellos están las clásicas noradrenalina y dopamina, además de la somatostatina, de la cual existen algunos análogos, como el octeotride.
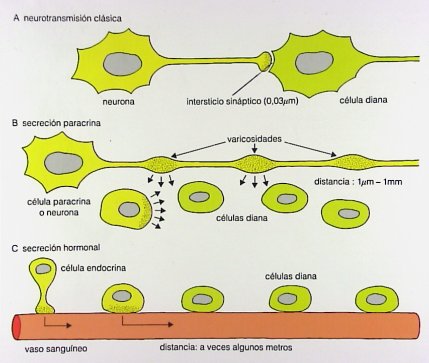
La transmisión de información se puede efectuar: mediante la neurotransmisión clásica, que tiene lugar en el intersticio sináptico, entre la neurona y la célula diana o blanco (en este caso, la célula muscular o miocito); mediante la secreción paracrina, en la cual la célula paracrina o neurona produce sustancias con efecto estimulador o inhibidor de la musculatura intestinal o gástrica; o mediante la secreción hormonal por parte de las células endocrinas del tubo digestivo, cuyos productos hormonales son transportados por los vasos a largas distancias, a veces varios metros, para actuar en la célula diana o enterocito. Un ejemplo de esto es la colecistoquinina, que actúa como neurotransmisor motor (Fig. 5).
Figura 5. Mecanismos de transmisión de la información en el tubo digestivo
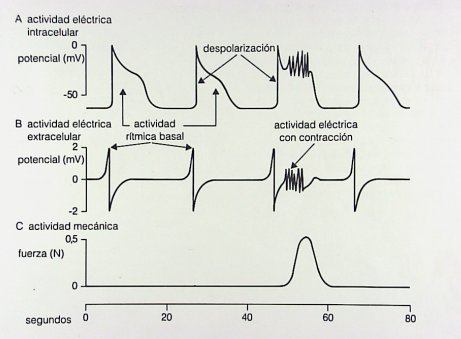
Figura 6. Actividades eléctrica y mecánica en las fibras de músculo liso del tubo digestivo
En el tubo digestivo ocurren, en general, dos tipos de contracciones: las contracciones fásicas, que son más cortas y rítmicas y se producen en el antro gástrico y en intestino delgado y las contracciones tónicas, que son más intensas y prolongadas y son propias del colon, como, por ejemplo, las contracciones masivas que causan la evacuación intestinal (Fig. 7).
Figura 7. Actividades eléctrica y mecánica en contracciones fásicas y tónicas
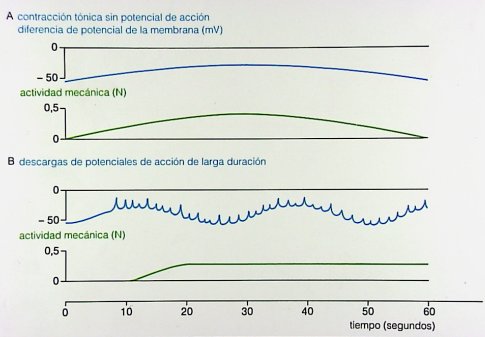
Según estudios publicados en los últimos ańos, esta actividad rítmica está controlada por el marcapasos compuesto por las células intersticiales de Cajal. Después de comer, el ritmo basal del estómago es de 3 oscilaciones por minuto, aumenta a alrededor de 8 a 10 en el duodeno y disminuye a 9 oscilaciones por minuto en el íleon. En general, la despolarización parcial no produce una contracción, mientras que la despolarización total se traduce en una contracción cuyas características dependerán del órgano en que se produzca. Las contracciones tónicas prolongadas del estómago ocurren en el fondo gástrico y no dependen de la descarga de potencial de membrana y son permanentes, es decir, el fondo gástrico tiene una contracción permanente, que no depende de potenciales de acción. En vesícula ocurre lo mismo: hay descargas de potenciales de larga duración relacionadas con las contracciones tónicas que ocurren, por ejemplo, en el vaciamiento del intestino grueso.
En la Fig 8 se ilustra el concepto general del reflejo peristáltico, que hace que el alimento se mueva de oral a caudal por contracción de las fibras musculares lisas circulares mientras se relajan las que quedan posteriores, lo que permite el avance del bolo alimenticio.
Figura 8. Avance del bolo digestivo por efecto del reflejo peristáltico
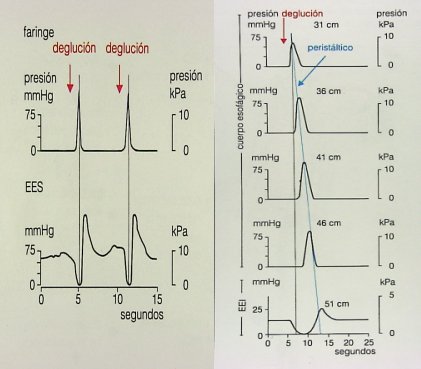
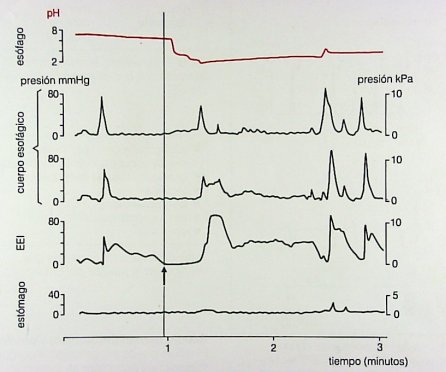
Cuando una persona se alimenta, la deglución se inicia con la contracción de la faringe y relajación del esfínter esofágico superior; una vez que el bolo alimenticio llega al cuerpo del esófago surge una onda transportadora, intensa, primaria, que persiste hasta que llega al final del esófago y el esfínter esofágico inferior se relaja. Esta situación corresponde a la fisiología normal y se puede registrar por manometría (Fig. 9).
Figura 9. Actividad esofágica eléctrica normal durante la deglución
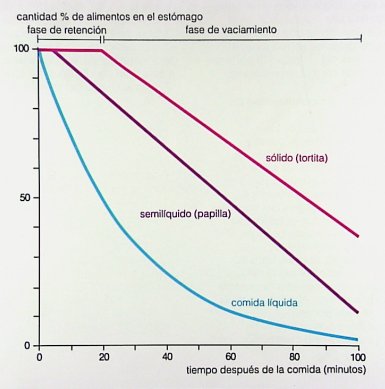
Cuando el bolo llega al estómago, en primer lugar ocurre una fase llamada de retención o de adaptación; luego la actividad cesa y da lugar al denominado íleo fisiológico, después del cual comienzan a ocurrir ondas fásicas, rítmicas, con una frecuencia de 3 contracciones por segundo, en cuerpo y antro. El tiempo de vaciamiento gástrico depende de la calidad del alimento: es más corto con comida líquida y más largo con sólidos o alimentos grasos y depende también de la cantidad ingerida (Fig. 10).
Figura 10. Tiempo de vaciamiento gástrico con diferentes tipos de alimentos
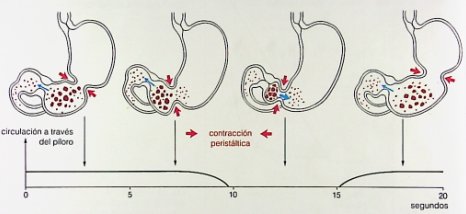
Un concepto importante en motilidad digestiva es que durante la fase gástrica postprandial también se producen relajaciones transitorias del esfínter esofágico inferior, que son normales y se presentan en número de 20 a 40 al día en los nińos y hasta 60 en adultos, sin constituir un proceso patológico; si las relajaciones transitorias del esfínter esofágico se exacerban, son muy repetidas o el clearance esofágico es deficiente, aparece el reflujo gastroesofágico patológico. Cuando el alimento está en el antro y cuerpo gástrico se producen ondas rítmicas que permiten el vaciamiento, la relajación del píloro y la consecuente entrada del alimento al intestino delgado (Fig. 11).
Figura 11. Ondas peristálticas que permiten el vaciamiento gástrico
Cuando el bolo alimenticio llega al duodeno también se producen ondas rítmicas fásicas determinadas por el marcapaso intestinal relacionado con las células de Cajal, con una frecuencia de 8 a 10 contracciones por minuto, que en el íleon aumenta a alrededor de 9 (Fig. 12).
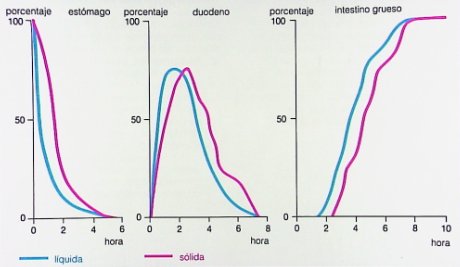
El vaciamiento gástrico puede demorar entre dos y cuatro horas, con diferencias entre líquidos y sólidos, diferencias que no son tan marcadas en duodeno e intestino grueso (Fig. 13).
Figura 12. Contracciones musculares, actividad eléctrica y presiones en estómago y duodeno, en relación con el vaciamiento
Figura 13. Tiempos transcurridos para vaciamiento gástrico, duodenal y de intestino grueso con diferentes tipos de alimentos
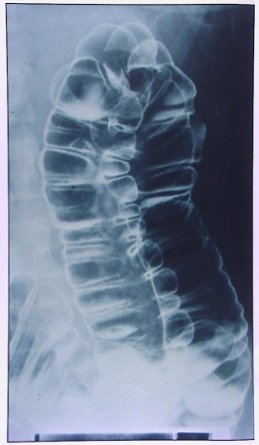
Una vez que llega el bolo alimenticio al intestino grueso sucede algo diferente: primero ocurren las llamadas contracciones haustrales, que también son rítmicas, seguidas por contracciones masivas, tónicas, que se presentan algunas veces en el día, en general después de comer y corresponden al reflejo gastrocólico. Las contracciones masivas, una vez que llegan al sigma, determinan que éste se contraiga lentamente y cause el acto de la defecación. La imagen muestra las haustras, que permiten que el alimento se mezcle y avance lentamente (Fig. 14).
Figura 14. Imagen radiológica de las haustras del intestino grueso.
Una vez que llega el bolo al colon transverso se producen contracciones masivas, que se transmiten a lo largo del colon hasta llegar a la zona del sigma, donde se producen las contracciones de defecación, proceso que puede ocurrir una o dos veces al día. Cuando la ingesta de fibra es baja, el bolo que llega al intestino grueso es duro y las haustraciones son de mala calidad, lo que determina un avance muy lento y constituye una de las causas de constipación funcional. En la zona del sigma, si el paciente retiene voluntariamente o rechaza el reflejo de defecación contrayendo el esfínter anal externo, puede ocurrir lo que en el concepto de Roma se llamaba constipación retentiva, con fecaloma y a veces escurrimiento.
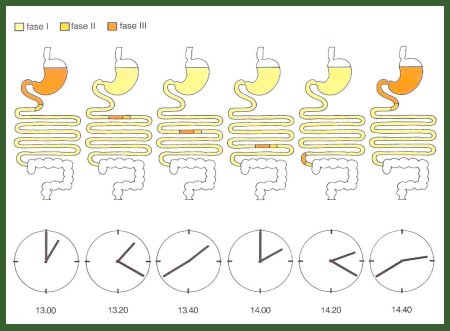
Otro concepto de gran relevancia en motilidad digestiva es el de complejo migratorio motor, que corresponde a lo que sucede después de que termina la digestión. Cuando termina el vaciamiento quedan residuos menores de dos milímetros de mucus y bacterias, tanto en el estómago como a lo largo de todo el intestino delgado, que son eliminados por el complejo migratorio motor. Éste tiene tres fases: la primera dura alrededor de 20 minutos y se caracteriza por reposo gástrico y ausencia de movimiento intestinal; en la segunda, el intestino delgado y el estómago comienzan a presentar contracciones tónicas lentas, determinadas por el marcapaso, similares a las que se observan después de la alimentación; por último, la fase de ayuno, que dura alrededor de una hora y cuarenta minutos, en que los residuos alimentarios llegan al intestino grueso (Fig. 15).
Figura 15. Complejo migratorio motor
Se ha visto que los nińos o personas que tienen este complejo alterado tienen mayor recuento bacteriano en el intestino delgado, lo que a veces constituye causa de diarrea. En el intestino grueso no hay complejo migratorio motor; las aves sí lo tienen y por eso eliminan deposiciones cada pocos minutos. En el ser humano no existe el complejo migratorio motor interdigestivo y por lo tanto no se presenta ese problema, que sería bastante desagradable.
En la segunda parte se presentará una reseńa de los principales trastornos de la motilidad digestiva en pediatría, su enfoque diagnóstico y las principales alternativas terapéuticas.
La edición y publicación de esta conferencia han sido posibles gracias al auspicio Nestlé Nutrition
Medwave Ańo VII, No. 7, Agosto 2007. Derechos Reservados.


 Esta obra de Medwave está bajo una licencia Creative Commons Atribución-NoComercial 3.0 Unported. Esta licencia permite el uso, distribución y reproducción del artículo en cualquier medio, siempre y cuando se otorgue el crédito correspondiente al autor del artículo y al medio en que se publica, en este caso, Medwave.
Esta obra de Medwave está bajo una licencia Creative Commons Atribución-NoComercial 3.0 Unported. Esta licencia permite el uso, distribución y reproducción del artículo en cualquier medio, siempre y cuando se otorgue el crédito correspondiente al autor del artículo y al medio en que se publica, en este caso, Medwave.
Este texto completo es la transcripción editada y revisada de una conferencia dictada en el Curso Desafíos y Oportunidades en Gastroenterología y Nutrición, organizado por la Sociedad Chilena de Pediatría durante los días 3, 4 y 5 de mayo de 2007. Directores: Dra. Sylvia Cruchet y Dr. Francisco Moraga.
Expositor:
Guillermo Venegas Villalobos[1]
Citación: Venegas G. Gastrointestinal motility: what is normal or abnormal? (I) - Anatomy and physiology of the digestive tract. Medwave 2007 Ago;7(7):e3245 doi: 10.5867/medwave.2007.07.3245
Fecha de publicación: 1/8/2007
Nos complace que usted tenga interés en comentar uno de nuestros artículos. Su comentario será publicado inmediatamente. No obstante, Medwave se reserva el derecho a eliminarlo posteriormente si la dirección editorial considera que su comentario es: ofensivo en algún sentido, irrelevante, trivial, contiene errores de lenguaje, contiene arengas políticas, obedece a fines comerciales, contiene datos de alguna persona en particular, o sugiere cambios en el manejo de pacientes que no hayan sido publicados previamente en alguna revista con revisión por pares.
Aún no hay comentarios en este artículo.
Para comentar debe iniciar sesión







 Estudios originales
Estudios originales