Revista Biomédica Revisada Por Pares

 Para Descargar PDF debe Abrir sesión.
Para Descargar PDF debe Abrir sesión.
Este texto completo es la transcripción editada y revisada de una conferencia dictada en el Curso Teórico y Seminarios de Oncología Básica, organizado por el Centro de Oncología Preventiva de la Universidad de Chile durante los días 3 de abril y 7 de agosto de 2002.
Editor Científico: Dr. José Manuel Ojeda.
Una diferencia fundamental entre la conducta de las células normales y las células tumorales, visualizada en cultivos celulares (1-5), es que las células normales se dividen con un número limitado de ciclos de replicación ( 40 a 60 ciclos ) (6) hasta la senescencia celular. En cambio las células del tumor tienen la habilidad de proliferar indefinidamente, es decir, son inmortales (7).
Hay evidencia experimental sustancial que el envejecimiento celular es dependiente de la división celular y que la expectativa de vida celular total, es moderada por el número de generaciones celulares, no por el tiempo cronológico (8,9). Esto significa que hay un proceso molecular que ocurre durante el crecimiento celular que culmina con la cesación de la división celular.
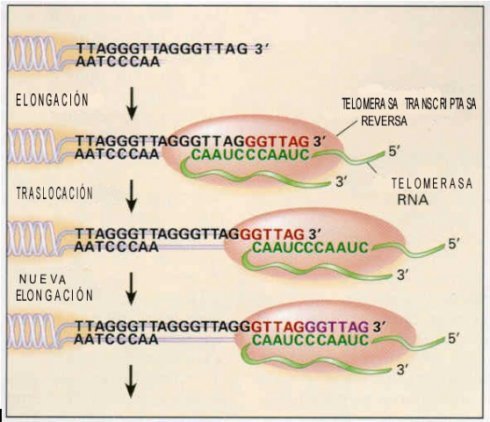
Un mecanismo que lleva a la muerte de una célula normal es la corrosión en cada división celular de la estructura en los extremos de los brazos del cromosoma, llamada TELÓMERO (del griego telos que significa extremo y meros que significa componente)(6).(Figura 1)
Antes de que cualquier célula pueda dividirse debe reproducir la doble cadena de ADN en sus cromosomas, pero tiene problemas al reproducir el ADN en los telómeros, donde hay más de 1000 secuencias cortas de bases TTAGGG repetidas una y otra vez junto a una variedad de proteínas que ligan ADN (6, 10,11).
Con cada división celular los telómeros se acortan en 50–200 pares de bases (12), principalmente porque en la síntesis de ADN la replicación de la cuerda 5`-3`se realiza sin problemas, sin embargo en la dirección 3`- 5` es incapaz de reproducir el extremo (6). Conocido como el “Problema de Repetición de Extremo” (13,14).
Figura 1. Visualización de los telómeros usando técnica de hibridación in situ con sondas fluorescentes. (FISH)
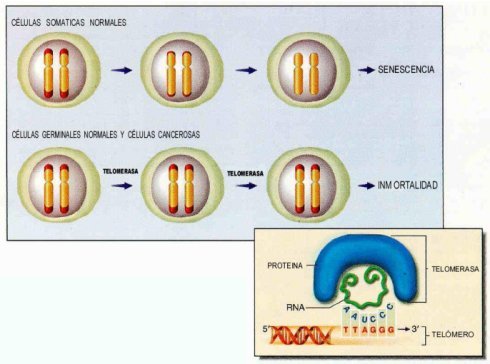
Es evidente que la pérdida progresiva del telómero del cromosoma es un mecanismo cronometrado importante en el envejecimiento celular humano. Cuando el telómero se ha puesto suficientemente corto, las células entran en una detención del crecimiento irreversible llamado senescencia celular, pudiendo ser este un potente mecanismo anticanceroso.
Los telómeros son las sucesiones de ADN repetitivas al final de los cromosomas lineales. En la mayoría de las células normales el telómero se va acortando progresivamente en cada ciclo celular. Cuando los telómeros son cortos, las células dejan de dividirse y sufren una detención del crecimiento (senescencia de la replicación).
Casi todas las células cancerígenas son inmortales, después de haber superado la senescencia celular reactivando la telomerasa–transcriptasa inversa que estabiliza los telómeros. Existe entonces, un complejo enzimático que mantiene los telómeros intactos, formada por la telomerasa, una ribonucleoproteína (RNP) de células eucarióticas y una transcriptasa inversa, conocida también como complejo telomerasa RNA-proteína (6,7,15,16).
El componente de ARN contiene nucleótidos que son complementarios a aquellos que presentan en el telómero. Por la transcripción inversa, la telomerasa hace una copia de ADN de su propia sucesión de ARN que se funde a los extremos 3´ al término del cromosoma.
La extensión de los telómeros por la telomerasa es totalmente opuesto a lo que ocurre normalmente con el encogimiento de los cromosomas después de cada ciclo de replicación de ADN.
La telomerasa humana sintetiza el tándem TTAGGG, repitiéndola sucesivamente al extremo 3´ de los cromosomas que usan parte de su mitad de ARN como templado (15).
El descubrimiento de la actividad de la telomerasa pone en estrecha correlación la inmortalización celular y la progresión maligna de los tumores (15) (Figura 2).
Figura 2.
Sin embargo, dos tipos de células, como son las germinales y las embrionarias, tienen actividad de telomerasa presente en forma habitual, ya que el cuerpo no se puede dar el lujo de perderlas. El resto de las células somáticas pierde la actividad de la telomerasa (6).
Acción de la telomerasa
Por la transcripción inversa , la telomerasa hace una copia de ADN de su propia sucesión de ARN ,
sintetizando el tandem TTAGGG, repitiéndola sucesivamente al extremo 3` al término de los cromosomas.
Por la transcripción inversa la telomerasa hace una hebra de ADN copiando su propia sucesión de ARN que se funde en los extremos 3´ al término del cromosoma. La extensión de los telómeros por la telomerasa se exige oponerse al encogimiento normal que sufría después de cada ciclo. La replicación normal de una hebra de ADN lineal por la ADN polimerasa sólo puede proceder de 5` a 3`. La longitud de un telómero está determinada entonces, por el equilibrio entre el número de divisiones celulares y la actividad de la telomerasa (16).
En los humanos las células germinales mantienen la longitud del telómero por acción de la telomerasa, a lo largo de la vida. En contraste los tejidos somáticos no tienen la telomerasa activa y pierden la longitud del telómero progresivamente y es así como los telómeros de las células de la piel y de la sangre son más cortos que aquellos de las células germinales, con una pérdida estimada de 15 a 40 nucleótidos por ańo (17,18).
Estas observaciones han llevado a la hipótesis cómo la longitud del telómero sirve de reloj biológico para las células normales (16). (Figura 3)
Figura 3. Estado de los telómeros durante el envejecimiento e inmortalización celular.
La expresión aberrante de la telomerasa en las células cancerígenas puede ser resultado de una mutación en las sucesiones que normalmente regulan su expresión y el tiempo de esa mutación puede inferirse de la longitud del propio telómero.
Se encontraron tumores ováricos que expresan la telomerasa y tienen telómeros muy cortos, haciendo pensar que ellos habían sufrido muchas divisiones celulares antes de que se activara la telomerasa y la longitud del telómero se estabilizara. Así, la activación del telómero no puede ser el evento transformador inicial que permita al cáncer progresar confiriendo inmortalidad a las células ya transformadas (19).
La mayoría de los cánceres humanos tiene telómeros cortos y niveles altos de telomerasa, considerando que en la mayoría, la telomerasa de los tejidos somáticos normales está ausente (20,21).
La telomerasa se ha examinado en los centenares de estudios como un biomarcador para diagnóstico precoz, pronóstico o estudio de enfermedad residual de un cáncer (22-29).
Los métodos descubiertos para la medición de la actividad telomerasa, usados en los ensayos de investigación, están disponibles comercialmente para evaluar diferente tipos de tejidos como biopsias de tumor congeladas, frescas, fluidos y secreciones.
Dos problemas centrales están determinando: cómo la longitud corta del telómero seńala la entrada a la fase senescente en el ciclo replicativo de las células normales y cómo la longitud del telómero es mantenida por la telomerasa en las células tumorales (7).
Para contestar estas importantes preguntas se está investigando en dos áreas:
Los telómeros están ocultos de la maquinaria celular que trataría el extremo de una molécula de ADN lineal como una cuerda rota que necesita la reparación. El primer trabajo fue el realizado por el laboratorio de Lange (30-33) quienes identificaron dos de las mayores proteínas que componían los telómeros, las telomeric DNA binding protein y las denominaron telomeric repeat binding factor (TRF) 1 y 2 .
Ambas TRF1 y TRF2 se expresan en todos los tipos celulares humanos, se repiten a lo largo de todo el ciclo celular e influyen directamente o por interacción con otros factores en la regulación de la longitud del telómero (34,35).
La ribonucleoproteína (RNP) telomerasa humana (complejo telomerasa) está compuesta de una proteína catalítica (hTERT) o human telomerase reverse transcriptase y una hebra de RNA de 451 bp (hTR) o human telomerase RNA , ambas son esenciales para la actividad telomerasa (36,37).
El componente de la proteína telomerasa transcriptasa inversa (hTERT) se requiere para la producción de actividad de telomerasa. Por esto se han ideado técnicas de inmunohistoquímica de proteína hTERT en las células tanto normales como cancerígenas.
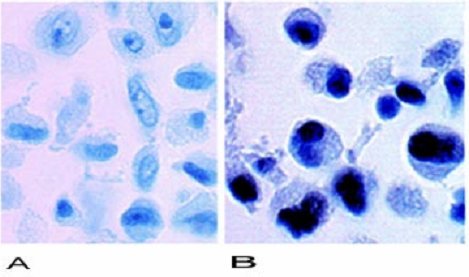
Es así, como en la siguiente fotografía podemos observar tinciones inmunohistoquímicas de células de cáncer como HeLa y HT1080 (de cultivos celulares ) y fibroblastos normales con cDNA de la hTERT introducidos, expresan niveles altos de la proteína telomerasa, pero esta no se observa en las células normales. Las células con actividad telomerasa tienen signos nucleares positivos, mientras que las sin actividad telomerasa no los tienen (Figura 4).
Figura 4. Técnicas inmunohistoquimicas para la detección de actividad telomerasa.
A. Células Normales (control)
B. Células Telomerasa Positivas.
En pediatría oncológica han aparecido una serie de trabajos que involucran la actividad de la telomerasa en el diagnóstico, evolución con quimioterapia y pronóstico de algunos tumores propios de la infancia.
En 1998, en el Memorial Sloan-Kettering Center de Nueva York, se evaluó la actividad de la telomerasa en 35 pacientes pediátricos con leucemia linfática aguda, leucemia mioeloide aguda y tumores sólidos pre y post quimioterapia.
Los resultados demostraron que en las leucemias, al diagnóstico la telomerasa estaba alta en muestra de médula ósea y sangre periférica y que disminuyó después de la inducción correlacionándose con la remisión. Los telómeros se fueron acortando como consecuencia de la quimioterapia con repetidos ciclos de regeneración hematopoyética. Por lo que se sugiere que la actividad de la telomerasa puede ser usada en el manejo de las malignidades infantiles (38).
Un estudio alemán publicado en 1999, realizado en 67 pacientes con neuroblastoma y dos ganglioneuromas, encontró la presencia de actividad telomerasa en un 20% de los neuroblastoma, en todas sus etapas y ausente en los ganglioneuromas.
Analizados sus datos encuentran una fuerte correlación estadística entre la presencia de actividad de telomerasa y mal pronóstico clínico en todos los estadios del tumor. Incluso su análisis de multivarianza revela la actividad de telomerasa como un marcador pronóstico independiente, que puede ser usado junto a otros marcadores como el n-myc para la evaluación final del paciente (39).
Siguiendo con sus estudios, este mismo grupo alemán en el 2000 publicó investigación con relación al impacto pronóstico de la presencia de actividad telomerasa y la expresión genética de sus subunidades, evaluadas por técnicas moleculares e inmunohistoquimicas, realizadas ya sea en tejidos frescos o fijados en parafina.
Los niveles de actividad telomerasa de 75 casos de neuroblastoma fueron correlacionados con la expresión génica de sus subunidades hTERT, hTR y TP1 realizado por RT-PCR.
La actividad telomerasa estuvo presente en el 29 % de los casos y se correlacionó significativamente con la sobrevida total y la sobrevida libre de eventos (p < 0.0001).
La significativa correlación con los resultados clínicos, hace que los autores recomienden la incorporación del análisis de la actividad telomerasa al estudio diagnóstico de rutina de estos tumores. Porque la mera presencia o ausencia de actividad telomerasa sin que sea cuantificada es suficiente para predecir el resultado de la enfermedad a largo plazo (40).
Una de las últimas publicaciones de la actividad telomerasa en pediatría, es un trabajo realizado en la Universidad de Virginia, Estados Unidos, donde se evaluó la telomerasa en la leucemia transitoria y en la leucemia aguda megacarioblástica ambas presentes en nińos con síndrome de Down, siendo la primera benigna y la otra maligna.
Los autores encontraron actividad de telomerasa en el 52 % de las leucemias megacarioblásticas y en el 12 % de las leucemias transitorias, la mayoría de estas últimas se asociaron a un desenlace fatal por la gravedad del cuadro. Observaron además, que la presencia de telomerasa puede ser un factor crítico en la malignización de células leucémicas (40).
La biología del telómero es importante en el cáncer humano. Las células malignas necesitan un mecanismo para mantener el telómero si ellas se van a dividir indefinidamente y la telomerasa resuelve ese problema. Aunque nosotros estamos empezando a identificar un número creciente de telómeros y las proteínas telomerasa asociadas, la clave es entender como el telómero, la telomerasa y su complejo enzimático actúan recíprocamente para mantener la longitud del telómero.
El desafío es aprender como intervenir en esos procesos y utilizar nuestro conocimiento creciente de biología del telómero para el diagnóstico y tratamiento de las enfermedades malignas.

 Esta obra de Medwave está bajo una licencia Creative Commons Atribución-NoComercial 3.0 Unported. Esta licencia permite el uso, distribución y reproducción del artículo en cualquier medio, siempre y cuando se otorgue el crédito correspondiente al autor del artículo y al medio en que se publica, en este caso, Medwave.
Esta obra de Medwave está bajo una licencia Creative Commons Atribución-NoComercial 3.0 Unported. Esta licencia permite el uso, distribución y reproducción del artículo en cualquier medio, siempre y cuando se otorgue el crédito correspondiente al autor del artículo y al medio en que se publica, en este caso, Medwave.
Este texto completo es la transcripción editada y revisada de una conferencia dictada en el Curso Teórico y Seminarios de Oncología Básica, organizado por el Centro de Oncología Preventiva de la Universidad de Chile durante los días 3 de abril y 7 de agosto de 2002.
Editor Científico: Dr. José Manuel Ojeda.
Autora:
Patricia Alvarez Aguayo[1]
Citación: Alvarez P. Telomerase in pediatric oncology. Medwave 2002 Nov;2(10):e3172 doi: 10.5867/medwave.2002.10.3172
Fecha de publicación: 1/11/2002
Nos complace que usted tenga interés en comentar uno de nuestros artículos. Su comentario será publicado inmediatamente. No obstante, Medwave se reserva el derecho a eliminarlo posteriormente si la dirección editorial considera que su comentario es: ofensivo en algún sentido, irrelevante, trivial, contiene errores de lenguaje, contiene arengas políticas, obedece a fines comerciales, contiene datos de alguna persona en particular, o sugiere cambios en el manejo de pacientes que no hayan sido publicados previamente en alguna revista con revisión por pares.
Aún no hay comentarios en este artículo.
Para comentar debe iniciar sesión
Medwave publica las vistas HTML y descargas PDF por artículo, junto con otras métricas de redes sociales.
Shay JW, Wright WE and Werbin H. Defining the molecular mechanisms of human cell immortalization. Biochim Biophys Acta. 1991 Apr 16;1072(1):1-7.
| PubMed |Hahn WC, Counter CM, Lundberg AS, Beijersbergen RL, Brooks MW and Weinberg RA. Creation of human tumour cells with defined genetic elements. Nature. 1999 Jul 29;400(6743):464-8. | CrossRef | PubMed |Vogelstein B and Kinzler KW. The multistep nature of cancer. Trends Genet. 1993 Apr;9(4):138-41. | CrossRef |Wright WE and Shay JW. Telomere dynamics in cancer progression and prevention: fundamental differences in human and mouse telomere biology. Nat Med. 2000 Aug;6(8):849-51.
| CrossRef | PubMed |Buys Charles HCM. Telomeres, telomerase and cancer. N Engl J Med. 2000 Apr 27;342(17):1282-3 | CrossRef | PubMed |Shay JW, Zou Y, Hiyama E and Wright WE. Telomerase and Cancer. Human Molecular Genetics, 10 (7), 677–685, 2001. | CrossRef | PubMed |Hayflick L. The limited in vitro lifetime of human diploid cell strains. Exp Cell Res. 1965 Mar;37:614-36.
| CrossRef |Shay JW and Wright WE. Haylick, his limit, and cellular aging Nat Rev Mol Cell Biol. 2000 Oct;1(1):72-6.
http://dx.doi.org/10.1038/35036093 | CrossRef | PubMed |Moyzis RK, Buckingham JM, Cram LS, Dani M, Deaven LL, Jones MD, et al A highly conserved repetitive DNA sequence, (TTAGGG)©ú, present at the telomeres of human chromosomes. Proc Natl Acad Sci U S A. 1988 Sep;85(18):6622-6. | CrossRef |Blackburn EH. The end of the (DNA) line. Nat Struct Biol. 2000 Oct;7(10):847-50.
| CrossRef | PubMed |Huffman KE, Levene SD, Tesmer VM, Shay JW, Wright WE. Telomere shortening is proportional to the size of the 3’ G – rich telomric overhang. J Biol Chem. 2000 Jun 30;275(26):19719-22.
| CrossRef | PubMed |Olovnikov AM. A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J Theor Biol. 1973 Sep 14;41(1):181-90.
| CrossRef |Gavory G, Balasubramanian Sh, Krupp G, Parwaresch R. PNA and Oligonucleotide Inhibitors of human Telomerase En: Telomerases, telomeres, and cáncer. EE.UU. Georgetown,2002. Haber DA. Telomeres,cancer, and inmortality. N Engl J Med. 1995 Apr 6;332(14):955-6.
| CrossRef | PubMed |de Lange T, Shiue L, Myers RM, Cox DR, Naylor SL, Killery AM,et al. Structure and variability of human chromosome ends. Mol Cell Biol. 1990 Feb;10(2):518-27. | PubMed | PMC |Hastie ND, Dempster M, Dunlop MG, Thompson AM, Green DK, Allshire RC, Telomere reduction in human colorectal carcinoma and with ageing. Nature. 1990 Aug 30;346(6287):866-8. | CrossRef | PubMed |Counter CM, Hirte HW, Bacchetti S, Harley CB. Telomerase activity in human ovarian carcinoma. Proc Natl Acad Sci U S A. 1994 Apr 12;91(8):2900-4. | CrossRef |Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL,et al. Specific association of human telomerase activity with inmortal cells and cancer. Science. 1994 Dec 23;266(5193):2011-5.
| CrossRef | PubMed |Shay JW, Bacchetti S. A survey of telomerase activity in human cancer. Eur J Cancer. 1997 Apr;33(5):787-91. | CrossRef |Breslow RA, Shay JW, Gazdar AF, Srivastava S. Telomerase and early detection of cancer : a national cancer institute workshop. J Natl Cancer Inst. 1997 May 7;89(9):618-23. | CrossRef |Shay JW. Telomerase in cancer : diagnostic, prognostic and therapeutic implications Cancer J Sci Am. 1998 May;4 Suppl 1:S26-34. | PubMed |Norton JC, Holt SE, Wright WE, Shay JW. Enhanced detection of telomerase activity in tumor derived human cell lines. DNA Cell Biol. 1998 Mar;17(3):217-9.
| CrossRef | PubMed |Hiyama E, Hiyama K, Yokoyama T, Matsuura Y, Piatyszek MA, Shay JW. Correlating telomerase activity levels with human neuroblastoma outcomes. Nat Med. 1995 Mar;1(3):249-55. | CrossRef | PubMed |Sommerfeld HJ, Meeker AK, Piatyszek MA, Bova GS, Shay JW, Coffey DS. Telomerase activity : A prevalent marker of malignant human prostate tissue. Cancer Res. 1996 Jan 1;56(1):218-22.
| PubMed |Mehle C, Piatyszek MA, Ljungberg B, Shay JW, Roos G. Telomerase activity in human renal cell carcinoma. Oncogene. 1996 Jul 4;13(1):161-6.
| PubMed |Tatsumoto N, Hiyama E, Murakami Y, Imamura Y, Shay JW, Matsura I,et al. High telomerase activity is an independent prognostic indicator of poor outcome in colorectal cancer. Clin Cancer Res. 2000 Jul;6(7):2696-701.
| PubMed |Shay JW, Gazdar AF. Telomerase in the early detection of cancer. J Clin Pathol. 1997 Feb;50(2):106-9. | CrossRef | PubMed | PMC |Chong L, van Steensel B, Broccoli D, Erdjument-Bromage H, Hanish J, A human telomeric protein. Science. 1995 Dec 8;270(5242):1663-7. | CrossRef | PubMed |Broccoli D, Smogorzewska A, Chong L, de Lange T. Human telomeres contain two distinct Myb – related protein, TRF1 and TRF2 . Nat Genet. 1997 Oct;17(2):231-5. | CrossRef | PubMed |van Steensel B, Smogorzewska A, de Lange T. TRF2 protects human telomeres from end–to–end fusions. Cell. 1998 Feb 6;92(3):401-13. | CrossRef |Bianchi A, Stansel RM, Fairall L, Griffith JD, Rhodes D, de Lange T. TRF1 binds a bipartite telomeric site with extreme spatial flexibility. EMBO J. 1999 Oct 15;18(20):5735-44. | CrossRef | PubMed | PMC |Smith S, Giriat L, Schmitt A, de Lange T. Tankyrase a poly (ADP-ribose) polymerase at human telomeres. Science. 1998 Nov 20;282(5393):1484-7. | CrossRef | PubMed |Smith S, de Lange T. Cell cycle dependent localization of the telomeric PARP, tankyrase, to nuclear pore complexes and centrosomes. J Cell Sci. 1999 Nov;112 (Pt 21):3649-56. | PubMed |Bodnar AG, Ouelette M, Frolkis M, Holt SE, Chiu CP, Morin GB,et al. Extension of life – span by introduction of telomerase into normal human cells. Science. 1998 Jan 16;279(5349):349-52. | CrossRef | PubMed |Weinrich SL, Pruzan R, Ma L, Ouelette M, Tesmer VM, Holt SE, et al. Reconstitution of human telomerase with the catalytic protein subunit hTRT. Nat Genet. 1997 Dec;17(4):498-502.
| CrossRef | PubMed |Engelhardt M, Ozkaynak MF, Drullinsky P, Sandoval C, Tugal O, Jayabose S,et al. Telomerase activity and telomere lengh in pediatric patients with malignancies undergoing chemotherapy. Leukemia. 1998 Jan;12(1):13-24.
| CrossRef | PubMed |Poremba C, Willenbring H, Hero B, Christiansen H, Schäfer KL, Brinkschmidt C,et al. Telomerase activity distinguishes between neuroblastomas with good and poor prognosis. Ann Oncol. 1999 Jun;10(6):715-21. | CrossRef | PubMed |Poremba C, Scheel C, Hero B, Christiansen H, Schafer KL, Nakayama J,et al. Telomerase activity and telomerase subunits gene expresion patterns in neuroblastoma : a molecular and immunohistochemical study establishing prognostic tools for fresh – frozen and paraffin – embedded tissues. J Clin Oncol. 2000 Jul;18(13):2582-92.
| PubMed |






 Estudios originales
Estudios originales